Эволюция дыхательной системы птиц. Эволюция дыхательной системы позвоночных. Онтофилогенетически обусловленные аномалии и пороки развития у человека
Методическая разработка урока биологии, предложенная на , включает подробный план-конспект занятия и презентацию на тему «Строение дыхательной системы позвоночных животных».
В ходе урока углубляются знания школьников об органах дыхания представителей животного мира. Осваивается и материал о системах органов, а также отдельных органах дыхания животных и их функциональные особенности.
Основные задачи, реализуемые в процессе , сформулированы следующим образом:
сформировать у учащихся представления о новых для них понятиях системы дыхания позвоночных животных;
развить теоретическое мышление, обеспечить усвоение приемов и методов логического преобразования информации;
воспитать у детей бережное отношение к окружающей их природе.
Строение дыхательной системы позвоночных животных — ход занятия
 Для успешной организации класса, в начале урока, автор методической разработки рекомендует написать с учениками пожелания на лепестках цветка и прикрепить на доску. После чего и знакомим класс с темой урока.
Для успешной организации класса, в начале урока, автор методической разработки рекомендует написать с учениками пожелания на лепестках цветка и прикрепить на доску. После чего и знакомим класс с темой урока.
Прежде чем приступить непосредственно к изучению новой темы, необходимо вспомнить пройденный материал, осуществив актуализацию ранее полученных знаний, вспомнив о дыхательной системе беспозвоночных животных. Для этого фронтально или по желанию учащиеся получают задания на карточках, где изображён живой организм одного из беспозвоночных животных. Требуется рассказать о дыхательной системе данного животного: какие у него имеются органы дыхания, а также кратко описать сам процесс дыхания . Ответы учащихся сопровождаются демонстрацией изображений животных на слайдах, где представлены: гидра, беззубка, червь, насекомое, паук и улитка.
Строение дыхательной системы у рыб
Вспоминаем с классом, каких животных принято называть позвоночными? Разделяем позвоночных на водных и наземных, исходя из среды их обитания. В свою очередь водные позвоночные представлены рыбами, а наземные позвоночные включают: земноводных, пресмыкающихся, млекопитающих и птиц.
Обращаем внимание школьников на экран. На слайде изображена рыба и строение её дыхательной системы, где отображены жаберные крышки и сами жабры. Из рисунка видно, что жабры включают в себя жаберные дуги и тончайшие жаберные лепестки, пронизанные крохотными кровеносными сосудами. При работе жабр, из воды поступает в кровь рыбы кислород, а из самой крови удаляется углекислый газ обратно в воду.
Строение дыхательной системы у земноводных
После изучения строения дыхательной системы рыб, переходим к рассмотрению особенностей системы дыхания земноводных. В качестве примера, из представителей земноводных будет рассмотрена дыхательная система лягушки в сопровождении также слайдов презентации. Здесь внимание обращаем на то, что легкие у земноводных расположены уже в грудной части организма.
Примечателен и следующий слайд, где демонстрируется картинка-схема, отражающая специфику вдоха лягушки, подробно описанную в разработке. Учитель также рассказывает и о строении легких, имеющих вид продолговатых мешочков, защищённых эластичными тонкими стенками. Внутренняя часть лёгких пронизана множественными разветвлёнными капиллярами. Однако земноводным присуще не только легочное, но и кожное дыхание. Так, через кожу земноводные способны дышать и на суше, и в воде.
О строении дыхательной системы пресмыкающихся
Дыхательную систему пресмыкающихся рассматриваем на примере ящерицы. Её легкие, подобно земноводным, имеют форму мешочков. Однако внутренняя поверхность этих мешочков значительно больше, нежели у земноводных, что связано с отсутствием кожного дыхания у пресмыкающихся.
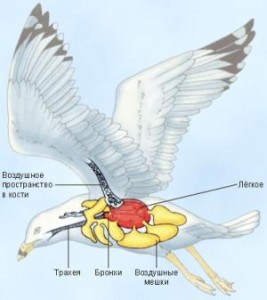
Следующими позвоночными животными, о дыхательной системе которых будет также рассмотрено на станут птицы. Сопровождая слайдами презентации, отмечаем наличие у птиц трахеи в дыхательной системе. А лёгкие птиц в сравнении с пресмыкающимися состоят из плотных губчатых тел и имеют множество ветвящихся полостей — бронхи. Также отличительным признаком является и наличие тонкостенных воздушных мешков в лёгких.
Строение дыхательной системы млекопитающих
 Приступаем на следующем этапе урока к изучению особенностей дыхательной системы млекопитающих, которые дышат атмосферным воздухом. Как и у предыдущих позвоночных, у млекопитающих органом дыхания являются лёгкие, отличающиеся ещё большей разветвлённостью бронхов. Вдох и выдох у млекопитающих происходит при чередующемся сокращении и расслаблении диафрагмы, а также межреберных мышц.
Приступаем на следующем этапе урока к изучению особенностей дыхательной системы млекопитающих, которые дышат атмосферным воздухом. Как и у предыдущих позвоночных, у млекопитающих органом дыхания являются лёгкие, отличающиеся ещё большей разветвлённостью бронхов. Вдох и выдох у млекопитающих происходит при чередующемся сокращении и расслаблении диафрагмы, а также межреберных мышц.
Закрепляется изученный материал при помощи дидактических карточек с вопросами по теме. И в заключение учитель предлагает учащимся ещё одно задание по таблице, где перечислены типы дыхания животных, а учащимся требуется прикрепить рисунок того или иного животного к соответствующему типу, учитывая изученные на уроке дыхательные системы позвоночных.
Подробную методическую разработку «Строение дыхательной системы позвоночных животных» к уроку биологии и презентацию можно скачать в начале статьи.
Глава 5.
Эволюция дыхательной системы.
5.1. Происхождение и функции дыхательной системы.
Впервые дыхательная система появляется у морских кольчатых червей - пескожила и нереиды, у которых на спинных ветвях параподий расположены примитивные жабры. Кроме того, у кольчатых червей газообмен происходит через богатую кровеносными сосудами кожу.
У млекопитающих и моллюсков строение органов дыхания зависит от условий их обитания: у водных форм - это жабры, способные использовать растворённый в воде кислород; у наземных - лёгкие и трахеи, приспособленные к использованию кислорода воздуха.
Так у ракообразных имеются жабры, которые расположены на ножках и ногочелюстях под боковыми складками головогрудного щита, где постоянно омываются водой.
Дыхательная система паукообразных представлена либо листовидными лёгкими, либо трахеями. Те и другие открывается наружу особыми отверстиями - стигмами на боковых частях члеников. В лёгочных мешках расположены многочисленные листовые складки, в которых проходят кровеносные капилляры. Лёгкие паукообразных гомологичны жабрам ракообразных. Трахеи представляют собой систему разветвленных трубочек, которые подходят непосредственно ко всем органам, где и совершается тканевой обмен.
У насекомых дыхание осуществляется с помощью трахей.
У многих членистоногих, имеющих тонкий хитиновый покров и относительно большую поверхность тела, наблюдается и диффузное дыхание.
У моллюсков в основном органами дыхания являются жабры за исключением наземных моллюсков, например, некоторых брюхоногих, которые утратили жабры, а их мантийная полость превратилась в легкое.
У многих беспозвоночных животных имеются приспособления, увеличивающие, дыхательную поверхность в виде местных специализированных органов дыхания.
5.3. Эволюция дыхательной системы у хордовых животных.
Дыхательная система всех хордовых животных и по происхождению, и топографически связана с кишечником, то есть с энтодермой.
У водных хордовых функцию дыхания выполняют жаберные щели, пронизывающие передний отдел кишечной трубки-глотки. У наземных хордовых жаберные щели закрываются во время эмбрионального развития, а затем исчезают. Функцию дыхания выполняют легкие, образующиеся из выпячивания кишечной трубки.
Эволюция жаберного аппарата у хордовых выражалось в уменьшении числа жаберных щелей при одновременном увеличении дыхательной поверхности путём образования жаберных лепестков.
Эволюция лёгких шла в направлении обособления дыхательных путей и увеличения дыхательной поверхности путем образования легких губчатого строения со сложной системой разветвления внутриклеточных бронхов, заканчивающихся пузырьками с ячеистыми клетками.
Наиболее примитивна дыхательная система у ланцетника, относящегося к низшим хордовым (подтип Бесчерепные). Передний отдел кишечника (стенка глотки) прободен жаберными щелями (до150 пар), которые открываются в артериальную (околожаберную) полость.
У круглоротых (подтип Позвоночные) органами дыхания являются также жаберные щели, но их уже меньше (5-15 пар). Они сообщаются с передним отделом кишечника и открываются наружу самостоятельными отверстьями.
Настоящие жабры появляются среди хордовых у рыб. Они представляют собой тонкие складки слизистой оболочки глотки, лежащие на жаберных дугах и снабжаемые венозной кровью через жаберные артерии, распадающиеся здесь на капилляры. У рыб имеется 4-7 (чаще5) жаберных мешков между жаберными дужками. Тычинки, расположенные на выпуклой поверхности жаберных дужек, препятствуют попаданию пищи из глотки в жабры.
Помимо жабр у рыб имеются добавочные органы дыхания, позволяющие им использовать кислород воздуха. Таким органом у рыб является плавательный пузырь. Стенки его богаты кровеносными сосудами, поэтому у некоторых зарывающихся в ил рыб он может служить для газообмена. Плавательный пузырь у большинства рыб развивается из дорзальных участков глотки и не является гомологом легких. Только у кистеперых рыб плавательный пузырь образуется как выпячивание вентральной части глотки и служит гомологом легких наземных животных, так как легкие позвоночных развиваются из брюшной части жаберного мешка.
У двоякодышащих рыб лёгкие есть, а плавательный пузырь отсутствует.
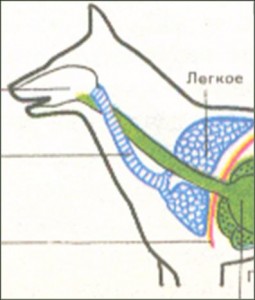
У личинок амфибий, как и у рыб, органы дыхания представлены древовидноветвящимися наружными жабрами. У большинства взрослых амфибий появляются лёгкие в виде тонкостенных парных выростов брюшной глотки позади последнего жаберного мешка. В связи с отсутствием грудной клетки и диафрагмы воздух в них попадает из ротовой полости за счет глотательных движений, осуществляющихся подбородочно-подъязычной мышцей. Материал жаберных дуг, следующий за подъязычной дугой, частично входит в состав хрящей гортани, которая появляется впервые у земноводных, являясь первым органом, относящимся к нижним дыхательным путям. Лёгкие начинаются непосредственно от гортани. Они крупноячеисты и имеют малую дыхательную поверхность, в связи с чем газообмен в большей степени осуществляется через кожные покровы, которые пронизаны большим количеством кровеносных капилляров и снабжены слизистыми железами.
У рептилий дыхательная система усложняется. Легкие у них уже мелко - ячеистые, они содержат многочисленные ячеистые перекладины и обладают большой дыхательной поверхностью. В дыхательных путях в связи с окончательным выходом рептилий на сушу наблюдается прогресс: выделяются верхние дыхательные пути, хотя и не окончательно отграниченные от ротовой полости - это носовая полость, а нижние – гортань, трахея и бронхи. Впервые появляется диафрагма, которая в дыхании принимает пассивное участие. Она либо частично разделяет грудную и брюшную полость, либо лишена мышечных волокон.
Механизм дыхания осуществляется за счет сокращения межреберных мышц, приводящих в движение грудную клетку.
У птиц легкие представляют собой плотно-губчатые тела пронизанные разветвлениями бронхов, а не мешки, как у рептилий. Кроме того, они дополняются в качестве резервуара для воздуха тонкостенными воздушными мешками. Последние располагаются между всеми воздушными органами, между мышцами, в полости кости и под костью.
При подъеме крыльев воздушные мешки через легкие наполняются воздухом, при опускании крыльев воздух через легкие выходит наружу. Таким образом, во время полета у птиц осуществляется двойное дыхание. Во время покоя птица дышит лишь путем расширения и сужения грудной клетки.
Органы дыхания у всех млекопитающих характеризуются сложностью как легких, имеющих альвеолярное строение, так и дыхательных путей. Идет дальнейшее по сравнению с предыдущими классами позвоночных животных разветвление бронхиального дерева. Трахея делиться на бронхи, которые ветвятся на бронхи второго, третьего и четвертого порядков и до самих мелких - бронхиол, на разветвлениях которых находятся альвеолы, легочные пузырьки имеющие в сумке огромную площадь(около 90см в квадрате), в которых происходит газообмен. Основная мышца, играющая важнейшую роль в акте дыхания, является диафрагма. Дыхательные пути млекопитающих выстланы мерцательным эпителием и полностью отделены от пищеварительной системы.
5.4. Аномалии и пороки развития дыхательной системы у человека.
1. В эмбриогенезе человека отражается первоначальная связь пищеварительной и дыхательной систем. В связи с этим становится понятным появление у человека большой группы врожденных пороков развития пищевода и трахеи типа эзофаготрахеальных свищей («жаберных щелей »).
2. Дизонтогенетические бронхолегочные кисты – округлая полоса в легких, отграниченная от окружающей ткани примитивно построенной стенкой недифференцированного бронха. Эти аномалии могут быть объяснены нарушением альвеолярной дифференцировки легочной ткани.
3. Кистозная гипоплазия – недоразвитие легкого. При этой патологии недоразвитыми оказываются целые доли легкого, которые представляют собой многочисленные полости, связанные с крупными бронхами и имеющие малую поверхность. Газообмен в таких участках легкого резко ослаблен.
4. Гипоплазия диафрагмы – недоразвитие диафрагмы от небольших дефектов в ее куполе до полной аплазии. Данная аномалия несовместима с жизнью и встречается чаще вместе с другими множественными пороками развития.
Глава 6.
Эволюция кровеносной системы.
6.1. Происхождение и функции кровеносной системы.
Функцией кровеносной системы является доставка ко всем органам тела кислорода и питательных веществ, удаления из организма продуктов распада и углекислоты, а также гуморальная функция.
Кровеносная система в основном имеет мезодермальное происхождение.
Эволюция кровеносной системы у беспозвоночных животных.
У низших беспозвоночных животных, т.е. у губок, кишечнополостных и плоских червей, доставка питательных веществ и кислорода от места их восприятия до частей тела происходит путем диффузных токов в тканевых жидкостях. Но у некоторых животных появляются пути, по которым идет циркуляция. Так возникают примитивные сосуды.
Дальнейшая эволюция кровеносной системы связана с развитием в стенках сосудов мышечной ткани, благодаря чему они могут сокращаться, а еще позже эволюция связана с превращением жидкости, заполняющей сосуды в особую ткань – кровь, в которой образуются различные кровяные клетки.
Кровеносная система бывает замкнутая и незамкнутая. Кровеносная система называется замкнутой, если кровь циркулирует только по сосудам, и незамкнутой, если сосуды открываются в щелевидные пространства полости тела, называемые синусами и лакунами.
Впервые кровеносная система появилась у кольчатых червей, она замкнутая. Имеется 2 сосуда – спинной и брюшной, связанные между собой кольцевыми сосудами, идущими вокруг пищевода. Движение крови происходит в определенном направлении – на спинной стороне к головному концу, на брюшной – назад благодаря сокращению спинного и кольцевых сосудов.
У членистоногих незамкнутая кровеносная система. На спинной стороне имеется пульсирующий сосуд, разделенный на отдельные камеры, так называемыми сердца, между которыми имеются клапаны. При последовательном сокращении сердец кровь поступает в сосуды, а затем изливается в щелевидные пространства между органами. Отдав питательные вещества, кровь медленно стекает в околосердечную сумку, а потом через парные отверстия в сердца.
У моллюсков кровеносная система также незамкнутая. Сердце состоит из нескольких предсердий, куда впадают вены и одного достаточно развитого желудочка, от которого отходят артерии.
Эволюция кровеносной системы у хордовых животных.
У низших хордовых, в частности у ланцетника, кровеносная система замкнутая, но сердца нет. Роль сердца выполняет брюшная аорта, от которой отходят приносящие жаберные артерии, в количестве 100-150 пар, несущие венозную кровь. Проходя через жаберные перегородки в неразветвленном виде кровь в артериях успевает окислиться и через выносящие парные жаберные артерии уже артериальная кровь поступает в корни спинной аорты, которые сливаются в непарную спинную аорту, от которой идут сосуды, несущие питательные вещества и кислород ко всем частям тела. Венозная кровь со спинной части собирается в передние и задние кардинальные вены, которые сливаются в левой и правой кювьеровы протоки, а из них в брюшную аорту. Кровь от брюшной стороны собирается в подкишечную вену, которая несет кровь в печень, где она обеззараживается, а оттуда по печеночной вене также впадает в кювьеров проток и далее брюшной сосуд.
У высших хордовых, в частности у низших позвоночных, т.е. у круглоротых и у рыб, усложнение кровеносной системы выражено в появлении сердца, которое имеет одно предсердие и один желудочек. В сердце бывает только венозная кровь. Круг кровообращения один, в котором артериальная и венозная кровь не смешиваются. Круговорот крови по телу сходен с кровеносной системой ланцетника. От сердца венозная кровь идет к жабрам, где окисляется, и от них окисленная (уже артериальная) кровь разносится по всему телу и по венам возвращается к сердцу.
С выходом животных на сушу и с появлением легочного дыхания, появляется второй круг кровообращения. Сердце получает не только венозную, но и артериальную кровь, и поэтому дальнейшая эволюция кровеносной системы идет по пути обособления двух кругов кровообращения. Это достигается делением сердца на камеры.
У земноводных и рептилий трехкамерное сердце, которое не обеспечивает полного разделения двух кругов кровообращения, поэтому еще происходит смешение артериальной и венозной крови. Правда, у рептилий желудочек уже разделен неполной перегородкой, а у крокодила четырех камерное сердце, поэтому смешение артериальной и венозной крови наблюдается в меньшей степени, чем у земноводных.
У птиц и млекопитающих сердце полностью разделено на четыре камеры – два предсердия и два желудочка. Два круга кровообращения, артериальная и венозная кровь не смешиваются.
Разберем эволюцию жаберных дуг у позвоночных животных.
У всех эмбрионов позвоночных животных впереди от сердца закладывается непарная брюшная аорта, от которой отходят жаберные дуги артерий. Они гомологичны артериальным дугам в кровеносной системе ланцетника. Но у них число артериальных дуг небольшое и равняется числу висцеральных дуг. Так у рыб их шесть. Первые две пары дуг у всех позвоночных испытывают редукцию, т.е. атрофируются. Оставшиеся четыре дуги ведут себя следующим образом.
У рыб разделяются на приносящие к жабрам и выносящие из жабер жаберные артерии.
Третья артериальная дуга у всех позвоночных, начиная с хвостатых амфибий, превращается в сонные артерии и несет кровь к голове.
Четвертая артериальная дуга достигает значительного развития. Из нее у всех позвоночных животных, опять же начиная с хвостатых амфибий, образуются собственно дуги аорты. У амфибий и рептилий парные, у птиц правая дуга (левая атрофируется), а у млекопитающих левая дуга аорты (правая атрофируется).
Пятая пара артериальных дуг у всех позвоночных, за исключением хвостатых амфибий, атрофируется.
Шестая пара артериальных дуг теряет связь со спинной аортой, из нее образуются легочные артерии.
Сосуд, связывающий во время зародышевого развития легочную артерию со спинной аортой, называется ботталовым протоком. Во взрослом состоянии он сохраняется у хвостатых амфибий и некоторых рептилий. Как результат нарушения нормального развития этот проток может сохранятся у других позвоночных и человека. Это будет врожденный порок сердца и необходимо в этом случае оперативное вмешательство.
Аномалии и пороки развития кровеносной системы у человека.
На основании изучения филогенеза сердечно-сосудистой системы становится понятным происхождения ряда аномалий и уродств у человека.
1. Шейная эктопия сердца – расположение сердца в области шеи. Сердце человека развивается из парных закладок мезодермы, которые сливаются и образуют единую трубку в области шеи. В процессе развития трубка смещается в левую часть грудной полости. Если сердце задерживается в области первоначальной закладки, то и возникает данный порок, при котором ребенок обычно погибает сразу после рождения.
2. Дестрокардия (гетеротопия) – расположение сердца справа.
3. Двухкамерное сердце – остановка развития сердца на этапе двух камер (гетерохрония). От сердца в этом случае отходит только один сосуд – артериальный ствол.
4. Незаращение первичной или вторичной межпредсердной перегородки (гетерохрония) в области овальной ямки, которая у эмбриона является отверстием, а также полное их отсутствие приводит к образованию трехкамерного сердца с одним общим предсердием (частота встречаемости 1:1000 рождений).
5. Незаращение межжелудочковой перегородки (гетерохрония) с частотой встречаемости 2,5-5:1000 рождений. Редким пороком является ее полное отсутствие.
6. Персистирование (нарушение дифференцировки) артериального, или ботталлова, протока , представляющего собой часть корня спинной аорты между 4-й и 6-й парами артерий слева. Когда легкие не функционируют, у человека во время зародышевого развития имеется ботталлов проток. После рождения проток зарастает. Сохранение его ведет к серьезным функциональным нарушениям, поскольку проходит смешанная венозная и артериальная кровь. Частота встречаемости 0,5-1,2:1000 рождений.
7. Правая дуга аорты – самая частая аномалия жаберных дуг артерий. При развитии происходит редукция левой дуги 4-й пары вместо правой.
8. Персистирование обеих дуг аорты 4-й пары, так называемое «Аортальное кольцо » - у эмбриона человека иногда не происходит редукции правой артерии 4-й жаберной дуги и корня аорты справа. В этом случае вместо одной дуги аорты развиваются две дуги, которые обогнув трахею и пищевод, соединяются в непарную спинную аорту. Трахея и пищевод оказываются в аортальном кольце, которое с возрастом сжимается. Порок проявляется нарушением глотания и удушением.
9. Персистирование первичного эмбрионального ствола . На определенной стадии развития у эмбриона имеется общий артериальный ствол, который потом разделяется спиральной перегородкой на аорту и легочной ствол. Если перегородка не развивается, то общий ствол сохраняется. Это приводит к смешению артериальной и венозной крови и обычно заканчивается смертью ребенка.
10. Транспозиция сосудов – нарушение дифференцировки первичного аортального ствола, при котором перегородка приобретает не спиральную, а прямую форму. В этом случае аорта будет отходить от правого желудочка, а легочной ствол – от левого. Этот порок встречается с частотой 1:2500 новорожденных и несовместим с жизнью.
11. Открытый сонный проток – сохранение комиссуры между 3-й и 4-й парами артериальных дуг (сонной артерией и дугой аорты). В результате увеличивается кровоток в мозг.
12. Персистирование двух верхних полых вен . У человека аномалией развития является наличие дополнительной верхней полой вены. Если обе вены впадают в правое предсердие, аномалия клинически не проявляется. При впадении левой вены в левое предсердие происходит сброс венозной крови в большой круг кровообращения. Иногда обе полые вены впадают в левое предсердие. Такой порок несовместим с жизнью. Данная аномалия встречается с частотой 1% от всех врожденных пороков сердечно-сосудистой системы.
13. Недоразвитие нижней полой вены – редкая аномалия, при которой отток крови от нижней части туловища и ног осуществляется через коллатерали непарной и полунепарной вен, являющихся рудиментами задних кардиальных вен. Редко встречается атрезия (отсутствие) нижней полой вены (кровоток осуществляется через непарные или верхнюю полую вену).
14. Отсутствие воротной системы печени .
Глава 7.
Эволюция выделительной системы.
7.1. Происхождение и функции выделительной системы.
Выделительная система имеет мезодермальное происхождение, а точнее образуется из нефротома сомитов. Она выполняет функцию удаления жидких продуктов обмена веществ из организма.
Эволюция выделительной системы у беспозвоночных животных.
Выделительная система в эволюции живых организмов претерпела большие изменения.
Если рассматривать группы животных в порядке усложнения их организации, то мы видим, что у кишечнополостных, тело которых состоит всего из двух слоев, обособленных органов выделения еще нет, а продукты обмена выделяются во внешнюю среду всей поверхностью тела.
Впервые выделительная система как самостоятельная система появилась у плоских червей в виде протонефридиев (греч. protos – первая, первичная, nephros – почка). Протонефридии начинаются в глубине паренхимы клетками звездчатой формы, получившими название конечных или терминальных клеток с мерцательным пламенем.
Эти клетки очень многочисленны, разбросаны они по всему телу и путем пиноцитоза отсасывают из тканевой жидкости паренхимы продукты обмена. В терминальных клетках имеются канальцы с пучком ресничек, которые своими колебаниями проталкивают продукты обмена сначала в канальцы с узким просветом, затем в каналы большей ширины, затем в главный выделительный канал (или каналы) и к выделительным порам.
У круглых червей выделительная система представляет 1-2 одноклеточными кожными железами, заменившими протонефридии. От желез отходят выросты в виде двух боковых каналов, лежащих в боковых валиках гиподермы. Сзади каналы оканчиваются слепо, а в передней части соединяются в один непарный канал, открывающийся наружу порой позади «губ». Функцией выделения обладают также и фагоцитарные клетки, расположенные по ходу выделительных каналов. В клетках накапливаются нерастворимые продукты распада, а также инородные тела, попадающие в полость тела.
Другой тип выделительной системы появляется у кольчатых червей, имеющих гомономную сегментацию и вторичную полость тела (целом). Это метамерно повторяющаяся в каждом сегменте пара метанефридиев, каждая из которых состоит из воронки, открывающейся в целом одного сегмента, называемая нефростомом, отходящего от него канальца, прободающего дессипимент и выделительной поры (нефропора) в другом сегменте.
У моллюсков и членистоногих вторичная полость редуцируется, и нефридии выделения значительно меняются. У ракообразных и большинства моллюсков они образуют компактные органы, напоминающие почки позвоночных. У ракообразных и насекомых вместо нефридиев появляются выделительные трубки, которые начинаются в миксоцеле и впадают в кишечник на границе среднего и заднего отделов. Они называются мальпигиевыми сосудами.
7.3. Эволюция выделительной системы у хордовых животных.
Выделительная система у низших хордовых построена по типу нефридиев. Так у ланцетника в области жаберных щелей метамерно расположено до 100 пар нефридиев, один конец которых открывается во вторичную, а другой – в околожаберную полость. Края целомического отверстия нефридиев (нефростом) имеют множество соленоцитов – клеток похожих на терминальные клетки протонефридиев. Следовательно, выделительные органы ланцетника имеют характер как прото-, так и метанефридиев.
У низших позвоночных (Anamnia) почки проходят две стадии: предпочки (головная или pronefros) и первичной (туловищная или mesonefros). У высших позвоночных (Amniota) развитие почек происходит в три стадии: предпочки, первичной и вторичной (тазовой или metanefros).
Структурной и функциональной единицей почек является нефрон, который также, как и почка, прошел длительный путь эволюции.
Почки закладываются, как уже уточнялось, в мезодерме, а именно в нефротомах. Выделительная система позвоночных связана с органами половой системы. Половые железы позвоночных, как правило, закладываются в виде парных складок на вентральной поверхности мезонефросов. Зачаток гонад состоит из утолщенного эпителия с большим количеством соединительной ткани.
Сначала мужские и женские половые железы имеют одинаковое строение. Позднее происходит их специализация и возникает связь различными для каждого вида частями выделительной системы, которые становятся половыми протоками.
У зародышей всех позвоночных животных закладывается головная почка или предпочка. Она состоит из 6-12 нефронов, продукты выделения которых собираются в общий мочеточник (парамезонефральный проток). Нефрон предпочки состоит из воронки (нефростома), которая выстлана ресничками и открывается в целом, и короткого прямого выделительного канала. Около воронок в стенках полости тела образуются грушевидные выросты из клубочков артериальных капилляров. Они отфильтровывают в целомическую полость как продукты выделения, так и полезные вещества. Целомическая жидкости попадает в воронки, канальцы и, собираясь в общий мочеточник, выводится в клоаку или мочевое отверстие. Несовершенство нефронов предпочки заключается в отсутствии прямой связи между кровеносной и выделительной системами, а также в постоянном присутствии в целомической жидкости продуктов выделения.
Предпочка во взрослом состоянии функционирует только у миксин (класс Круглоротые), а у всех остальных подвергается редукции (у зародыша человека она сохраняется около 40 часов).
У анамний после редукции предпочки появляется первичная почка.
Первичная почка закладывается в туловищных сегментах тела. Она содержит до нескольких сотен нефронов, продукты выделения которых собираются в выделительные протоки. Нефрон первичной почки состоит из: воронки (нефростома), которая выстлана ресничками и открывается в целом; почечного тельца, которое состоит из двустенной капсулы Боумена–Шумлянского и клубочка капилляров; извитого выделительного канала.
Продукты выделения из капилляров клубочка отфильтровываются в полость капсулы, по извитому канальцу собираются в мочеточник, мочевой пузырь и выводятся через клоаку или мочевые отверстия.
Нефрон первичной почки характеризуется рядом прогрессивных изменений:
появляется прямая связь между кровеносной и выделительной системами;
увеличивается количество нефронов в почке;
происходит удлинение и изменение формы извитого канальца, в результате чего начинают осуществляться процессы обратного всасывания нужных веществ, и происходит концентрация мочи;
уменьшается количество продуктов выделения в целоме.
У низших позвоночных (у миног из класса Круглоротые, у рыб и у амфибий) первичная почка функционирует в течении всей жизни как орган выделения.
У высших позвоночных (рептилий, птиц и млекопитающих) в том числе и человека первичная почка редуцируется.
У самок амниот часть канальцев первичной почки сохраняется в виде незначительных рудиментов epoophron и paraophron, а из остатков предпочки и мочеточника развивается яйцевод, который дифференцируется на отделы, а именно маточные трубы, матку и влагалище.
У самцов амниот пронефрос и ее мочеточник полностью редуцируются. Канальцы передней части первичной почки сохраняются и преобразуются в придатки семенника – эпидидимис, а мочеточник первичной почки превращается в семяпровод.
Основная роль первичной почки в эмбриогенезе – инициация образования вторичной почки.
Вторичная почка закладывается ниже первичной почки, но по мере роста и развития смещается кверху и, начиная с 3-го месяца, располагается выше первичной. Одна вторичная почка у человека содержит более миллиона нефронов.
Продукты выделения из вторичной почки собираются в мочеточники
Нефрон вторичной почки состоит из:
почечного тельца в капсуле Боумена – Шумлянского;
выделительного канальца, который дифференцируется на проксимальный, дистальный отделы и петлю нефрона (петля Генле).
Продукты выделения поступают в нефрон путем фильтрации крови в капсулах. Образуется первичная моча, у человека она составляет 170-180 литров в сутки. В извитых почечных канальцах первичная моча концентрируется за счет реабсорбции – обратного всасывания нужных веществ и образование вторичной мочи. Вторичная моча (1,7-1,8 литра в сутки у взрослого человека) собирается в мочеточники. Они образуются из боковых выростов мочеточника первичной почки.
Таким образом, в эволюции животных можно выделить три типа выделительных систем: протонефридии, метанефридии , почки. Эволюция этой системы в ряду позвоночных идет в направлении, во-первых, увеличения более тесной связи с кровеносной системой, во-вторых, увеличения выделительной поверхности за счет увеличения числа нефронов и, в-третьих, совершенствования строения самого нефрона, который теряет связь с целомической полостью, удлиняет почечный каналец и создает механизм обратного всасывания.
Процесс формирования мочеполовой системы позвоночных является ярким примером субстиции органов.
Субстиция – способ преобразования органов, при котором более ранние закладки органа после появления последующих редуцируются.
Аномалии и пороки развития выделительной системы у человека.
Сложность формирования почек делает возможность появления отклонений от нормального процесса. Аномалии развития почек бывают различными.
1. «Опущение почки» - одна из почек может не подняться из области таза в поясничную область и остаться там, где закладывалась, т.е. в области таза.
2. «Подковообразная почка» - при низком положении почек и срастании их нижними полюсами получается подковообразная почка.
3. Образование общей почечной массы – нарушение, при котором обе почки могут оказаться по одну сторону от средней линии и срастись в общую почечную массу.
4. Наличие третьей почки – число почек может быть больше или меньше нормального. Очень редко встречается третья почка, лежащая на позвоночном столб между двумя или ниже какой-либо из них.
5. Арения – врожденное отсутствие обеих почек, часто сочетается с другими врожденными пороками. Дети нежизнеспособны.
6. Агенезия почек – врожденное отсутствие одной из почек.
7. Гипоплазия почек – врожденное уменьшение массы и объема почек, может быть одно- и двусторонним. При односторонней гипоплазии наблюдается гипертрофия второй нормальной почки.
8. Поликистоз почек – двустороннее увеличение почек с формированием кист. Различают крупнокистозные и мелкокистозные почки.
Крупнокистозные почки характеризуются образованием в корковом слое большого числа крупных кист с прозрачным содержимым, между кистами сохраняются участки нормальной почечной паренхимы. С такими почками больные могут доживать до 40-50 лет.
9. Агенезия (отсутствие), атрезия (врожденное отсутствие или аномальное строение), стеноз (сужение просвета), эктопия (ненормальное расположение) устьев лоханок и мочеточников.
10. Удвоение мочеточников – частичное расщепление мочеточника или полное – два мочеточника с двумя лоханками и двумя устьями в мочевом пузыре.
11. Агенезия (отсутствие) мочевого пузыря.
12. Полное или неполное удвоение мочевого пузыря – при удвоении мочевого пузыря продольная перегородка делит пузырь на две самостоятельные части. Каждая часть пузыря имеет либо самостоятельное сообщение с отдельным для каждой из них мочеиспускательным каналом (т.е. имеется двойная уретра), или обе половины пузыря открываются в одну уретру, либо открывается в уретру только одна половина пузыря.
13. Расщепление мочеиспускательного канала – это нарушение может располагаться на его нижней (гипоспадия) или верхней (эписпадия) поверхности. Все пороки мочевыводящих путей ведут к нарушению оттока мочи и к появлению осложнений (гидронефроз, хронический пиелонефрит и др.). необходимо хирургическое вмешательство.
14. Эписпадия – врожденная аномалия, при которой нарушенное отверстие уретры находится на верхней поверхности полового члена, встречается очень редко.
ч. 1 ч. 2 ч. 3 ч. 4
Водные беспозвоночные. Обитатели водной среды - колыбели жизни на нашей планете - получают растворенный кислород непосредственно из воды, в воду же отдают углекислый газ, образующийся в ходе биологического окисления. Прямое дыхание свойственно одноклеточным - простейшим (как, впрочем, и некоторым многоклеточным организмам, например личинкам насекомых). Диффузия респираторных газов через всю поверхность тела присуща губкам, плоским червям.
Конвективный перенос газов появляется у иглокожих; у них наблюдается побуждаемый движением ресничек ток жидкости (где и растворяются газы) через полемическую полость. Здесь функция дыхания еще не отделена от питания и выделения.
В процессе дальнейшего развития «внешний» конвективный транспорт газов дополняется «внутренним»: возникает кровеносная система. Впервые замкнутый контур циркуляции появляется у немертин: их кровь уже содержит химический переносчик кислорода - гемоглобин. Вообще же в кислородном транспорте участвуют различные пигменты (хлоркруорины, гемеретрины, гемоцианины и гемоглобины), встречающиеся у некоторых представителей ряда типов животных: круглых, плоских и кольчатых червей, моллюсков, иглокожих, членистоногих.
Наземные беспозвоночные. Первые воздуходышащие многоклеточные появились в силурийский период. Выходу животных на сушу способствовало в числе прочих факторов обеднение водоемов кислородом при одновременном увеличении в атмосфере его парциального давления, которое в то время (около 450 млн. лет назад) было существенно ниже нынешнего. Молекулярный кислород в воздушной среде значительно доступнее, нежели в водной: скорость диффузии O 2 в воздухе примерно в 300 тыс. раз выше, чем в воде. Правда, поглощение этого газа организмом обязательно проходит стадию растворения в жидкости, покрывающей газообменные поверхности. Этот принцип сохраняется на всех этапах эволюции.
Первый из «сухопутных» типов дыхания встречается у кольчатых червей. Он осуществляется без посредства каких-либо респираторных органов. Газообмен происходит через увлажненный кожный покров, состоящий из однослойного эпителия и обильно снабженный кровеносными сосудами. Мышечные сокращения активно проталкивают кровь, содержащую гемоглобин, в глубокие ткани.
Другой тип обмена газов со средой отмечается у брюхоногих моллюсков. Здесь появляются уже специализированные органы дыхания - примитивные жабры в виде постоянно увлажненных лепестков либо мантийная полость, открывающаяся наружу отверстиями - пневмостомами. Незамкнутая кровеносная система обеспечивает с участием гемоглобина перенос кислорода к тканям.
Разнообразные формы дыхания встречаются у членистоногих. Предки этих животных обитали в воде, а многие (большая часть ракообразных) так и остались воднодышащими. С переходом на сушу листовидные жабры, связанные с конечностями, были преобразованы в мешковидные структуры, так называемые диффузионные легкие. Кислород из этих легких доставляется тканям через незамкнутую кровеносную систему с током гемолимфы, проталкиваемой трубчатым сердцем. Но у большинства членистоногих сформировались совсем новые органы дыхания - трахеи, проникающие глубоко в тело животного. Именно трахейный способ обмена газов характерен для насекомых.
|
Рис. 10.2 Трахейная система насекомого 1 - трахеи, ведущие к крыльям, 2 - дыхальца, 3 - трахеальные стволы. |
|
|
|
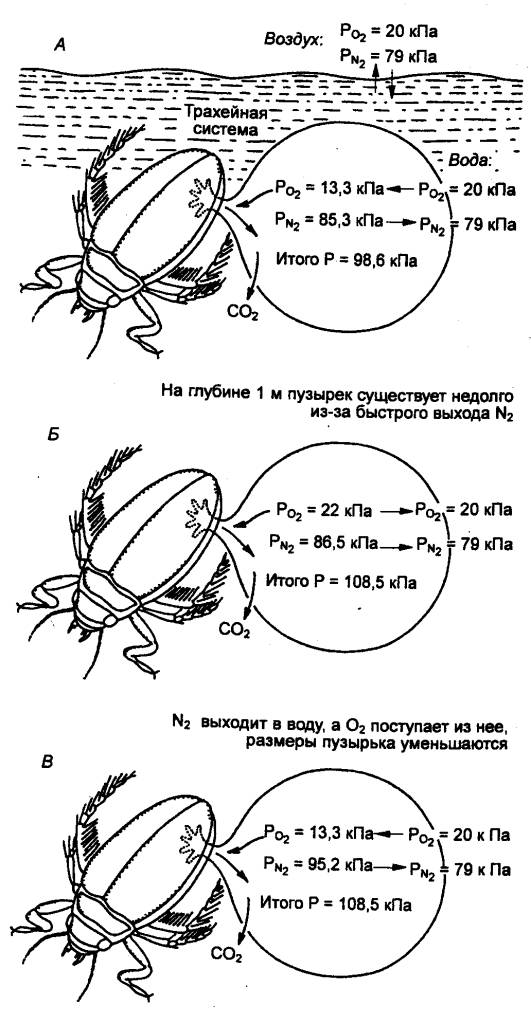
Рис. 10.3Парциальные давления О 2 , СО 2 и N 2 , а также суммарное давление (Р) в пузырьке, захваченном водным насекомым при нырянии А- состояние в начале погружения, Б - после быстрого погружения на глубину 1 м, В - состояние на той же глубине спустя некоторое время. Стрелками показано движение молекул газа. |
Развитие системы трахей у насекомых было обусловлено, с одной стороны, наличием хитинового покрова, а с другой - очень высокими в расчете на единицу массы тела энерготратами и, следовательно, интенсивным потреблением кислорода (в полете оно увеличивается в десятки, а то и в сотни раз). Дыхательная система насекомого (рис. 10.2) сообщается с наружным воздухом посредством дыхалец (стигм). Каждое дыхальце ведет в полость и далее в широко ветвящиеся трубочки, трахеи, делящиеся в основном в соответствии с сегментами тела. Это ветвление завершают узкие короткие трахеолы, которые достигают клеточных мембран или даже проникают внутрь клеток до митохондрий.
Основным механизмом трахейного дыхания служит диффузия газов, поддерживаемая разностью парциальных давлений между клетками и атмосферой. Кроме того, транспорт респираторных газов обеспечивается конвективными потоками, хотя и весьма непостоянными. Движение воздуха в трахеях вызывается ритмическими сокращениями туловища, которые управляются центрами, расположенными в ряде ганглиев нервной системы насекомого. Ритмические изменения объема груди в полете тоже содействуют вентиляции трахейной системы. Помимо того, дыхание регулируется в зависимости от потребностей тканей в кислороде открытием и закрытием дыхалец, а также активным перемещением кончиков трахеол и содержащейся в них жидкости - плазмы гемолимфы (сама гемолимфа существенной роли в газообмене не играет).
Трахейное дыхание позволяет насекомому приспосабливаться к самым различным условиям жизни. Вместе с тем лежащий в основе данного типа газообмена механизм диффузионного транспорта ограничивает расстояние, на которое переносится кислород. Это одна из причин, препятствующих представителям самого многочисленного класса животных достичь сколько-нибудь значительных размеров.
Некоторые водные насекомые (например, личинки москитов) дышат через водоотталкивающий сифон, выходящий из воды. Многие же при погружении захватывают с собой пузырьки воздуха. Так, жук - окаймленный плавунец ( Dytiscus marginalise ныряет с пузырьками воздуха под крыльями или у заднего конца брюшка (рис. 10.3). При погружении насекомого происходит обмен газами между пузырьком и тканями через трахейную систему. Помимо того, может происходить обмен газами между пузырьком и водой, причем скорость этого процесса зависит от градиентов парциальных давлений (напряжений) газов и площади поверхности раздела «вода-воздух».
Известно два типа дыхания: водное и воздушное.
У водных хордовых животных дыхание жаберное. Эволюция шла в сторону уменьшения количества жаберных щелей. Так у ланцетника в стенках глотки имеются 100-150 пар жаберных щелей.
У рыб - число жаберных перегородок сокращается до 5 пар, но увеличивается число жаберных лепестков. Жабры образуются из впячиваний глотки в виде жаберных мешков, а затем появляются перегородки с лепестками.
У земноводных появляется воздушное дыхание. Это мешковидные легкие, аналогичные плавательному пузырю кистеперых. Стенки мешков гладкие, тонкие, с небольшими перегородками. Воздухоносные пути отсутствуют. Дыхание у земноводных кожно-легочное.
У рептилий - кожа не выполняет дыхательную функцию. Легкие ячеистые появляются разветвленные перегородки. Появление воздухоносных путей. В трахее формируются хрящевые кольца, разделяясь, образуются два бронха.
У птиц - губчатые легкие , пронизанные бронхами. С легкими связано 5 пар воздушных мешков - тонкостенных, легко растяжимых выростов вентральных ответвлений крупных бронхов, находящиеся среди внутренних органов, между мышцами и в полостях трубчатых костей крыльев. Эти мешки играют большую роль в процессе дыхания птиц во время полета. Наряду с функцией дыхания воздушные мешки несут добавочные функции: облегчают массу тела птицы. При вдохе только 25 % наружного воздуха остаётся непосредственно в лёгких, а 75 % проходит через них и попадает в специальные воздушные мешки. При выдохе воздух из воздушных мешков опять идёт через лёгкие, но уже наружу, образуя так называемое двойное дыхание. Таким образом, лёгкие постоянно насыщаются кислородом как во время вдоха, так и выдоха.
У млекопитающих лёгкие альвеолярного типа. Формируется бронхиальное дерево, на концах бронхов - альвеолы. Появляется диафрагма , которая разделяет полость тела на грудную и брюшную.
Основные направления эволюции:
1. Смена жаберного дыхания на кожно-легочное, а затем - легочное.
2. Увеличение дыхательной поверхности легкого.
3. Появление и дифференцировка дыхательных путей.
Пороки развития дыхательной системы:
1. Сохранение жаберных щелей (жаберные свищи).
2. Атрезия трахеи.
3. Трахейно-пищеводные фистулы.
4. Агенезия (отсутствие) или гипоплазия (недоразвитие) доли или целого легкого.
5. Недоразвитие легкого, бронхов.
6. Добавочные доли или целое легкое.
7. Зеркальная закладка левого и правого легкого
Эволюция нервной системы позвоночных. Онтофилогенетически обусловленные аномалии и пороки развития у человека.
Нервная система осуществляет быстрое реагирование организма на изменение условий посредством рефлексов. Функции нервной системы: 1) Регуляторная; 2) координирующая; 3) связь с внешней средой; 4) интегрирующая, лежит в основе высшей нервной деятельности, поведенческих реакций, абстрактного мышления. Происхождение – эктодермальное. Это производная нервной трубки с невроцелем внутри. Передний отдел нервной трубки увеличивается в размерах, дифференцируется на отделы и преобразуется в головной мозг. Образование головного мозга – цефализация (кефализация). Головной мозг позвоночных закладывается из трех мозговых пузырей: переднего, среднего и заднего. Позже дифференцируется на 5 отделов: передний, промежуточный, средний, задний и продолговатый. Во всех отделах головного мозга различают мантию, или крышу, располагающуюся над желудочками, и основание, или дно, лежит под желудочками (желудочки – расширения невроцеля).
Аномалии и пороки
Большинство не совместимо с жизнью.
1. Рахисхис, или платиневрия, - отсутствие замыкания нервной трубки. Не совместим с жизнью.
2. Голопрозэнцефалия – полушария не разделены, кора недоразвита. Не совместим с жизнью.
3. Анэнцефалия – отсутствие переднего мозга.
4. Нарушение дифференцировки коры. Большинство умирают в течении 1 года жизни.
4.1. Агирия – отсутстивие извилин.
4.2. Олигогирия – малое количество извилин.
4.3. Пахигирия – утолщение извилин.
Основные направления эволюции нервной системы:
· Дифференцировка нервной трубки на головной и спинной мозг.
· Прогрессивное развитие переднего конца нервной трубки в головной мозг.
· Дифференцировка на отделы.
· Изменение системы развития отделов.
· Смещение центров регуляции нервной деятельности от среднего к ведущему переднему.
· Образование коры и высших чувствительных и двигательных центров психической деятельности.
· Увеличение площади головного мозга за счет появления извилин.
· Формирование функциональной ассиметрии полушарий.
· Увеличение черепно-мозговых нервов.
Эволюция головного мозга.
Формирование головного мозга у всех позвоночных начинается с образования на переднем конце нервной трубки трех вздутий или мозговых пузырей: переднего, среднего и заднего . В дальнейшем передний мозговой пузырь делится поперечной перетяжкой на два отдела. Первый из них образует передний отдел головного мозга , который у большинства позвоночных образует полушария большого мозга . На задней части переднего мозгового пузыря развивается промежуточный мозг. Средний мозговой пузырь не делится и целиком преобразуется в средний мозг. Задний мозговой пузырь также подразделяется на два отдела: в передней его части образуется задний мозг или мозжечок , а из заднего отдела образуется продолговатый мозг , который без резкой границы переходит в спинной мозг.
В процессе образования пяти мозговых пузырей полость нервной трубки образует ряд расширений, которые носят название мозговых желудочков . Полость переднего мозга носит название боковых желудочков, промежуточного - третий желудочек, продолговатого мозга четвертый желудочек, среднего мозга - сильвиев канал, который соединяет 3-й и 4-й желудочки. Задний мозг полости не имеет.
В каждом отделе мозга различают крышу, или мантию и дно, или основание. Крышу составляют части мозга, лежащие над желудочками, а дно - под желудочками.
Вещество мозга неоднородно. Темные участки - серое вещество, светлые - белое вещество. Белое вещество - скопление отростков нервных клеток с миелиновой оболочкой (много липидов, которые придают беловатую окраску). Серое вещество - скопление нервных клеток между элементами нейроглии. Слой серого вещества на поверхности крыши любого отдела мозга носит название коры.
У всех позвоночных головной мозг состоит из пяти отделов, расположенных в одной и той же последовательности. Однако, степень их развития неодинакова у представителей различных классов. Эти различия обусловлены филогенезом.
Выделяют три типа головного мозга: ихтиопсидный, зауропсидный и маммальный.
К ихтипсидному типу мозга относят мозг рыб и амфибий. Он является ведущим отделом головного мозга, центром рефлекторной деятельности.
· Головной мозг рыб имеет примитивное строение, что выражается в незначительных размерах мозга в целом и слабом развитии переднего отдела.
Передний мозг мал и не разделен на полушария. Крыша переднего мозга тонкая. У костистых рыб не содержит нервной ткани. Основную массу его образует дно, где нервные клетки образуют два скопления - полосатые тела. От переднего мозга вперед отходят две обонятельные доли. Передний мозг рыб выполняет функцию обонятельного центра.
Промежуточный мозг рыб сверху прикрыт передним и средним. От его крыши отходит вырост - эпифиз, от дна - воронка с прилегающим к ней гипофизом и зрительные нервы.
Средний мозг - наиболее развитый отдел мозга рыб. Это зрительный центр рыб, состоит из двух зрительных долей. На поверхности крыши находится слой серого вещества (кора). Это высший отдел мозга рыб, поскольку сюда приходят сигналы от всех раздражителей и здесь вырабатываются ответные импульсы.
Мозжечок рыб развит хорошо, поскольку движения рыб отличаются разнообразием.
Продолговатый мозг у рыб обладает сильно развитыми висцеральными долями, связан с сильным развитием органов вкуса.
· Головной мозг амфибий имеет ряд прогрессивных изменений, что связано с переходом к жизни на суше, которые выражаются в увеличении общего объема мозга и развитии его переднего отдела. Одновременно происходит разделение переднего мозга на два полушария. Крыша переднего мозга состоит из нервной ткани. В основании переднего мозга лежат полосатые тела. Обонятельные доли резко ограничены от полушарий.
Передний мозг попрежнему имеет значение лишь обонятельного центра.
Промежуточный мозг хорошо виден сверху. Крыша его образует придаток - эпифиз, а дно - гипофиз.
Средний мозг меньше по размерам, чем у рыб. Полушария среднего мозга хорошо выражены и покрыты корой. Это ведущий отдел ЦНС, т.к. здесь происходит анализ полученной информации и выработка ответных импульсов. Он сохраняет значение зрительного центра.
Мозжечок развит слабо и имеет вид небольшого поперечного валика у переднего края ромбовидной ямки продолговатого мозга. Слабое развитие мозжечка соответствует простым движениям амфибий.
К зауропсидному типу мозга относят мозг пресмыкающихся и птиц.
· У рептилий наблюдается дальнейшее увеличение объема головного мозга.
Передний мозг становится наиболее крупным отделом. Он перестает быть только обонятельным центром и становится ведущим отделом ЦНС за счет дна, где развиты полосатые тела. На поверхности мозга впервые в процессе эволюции появляются нервные клетки или кора, которая имеет примитивное строение (трехслойная) и получила название древней коры - археокортекс.
Промежуточный мозг сверху прикрыт полушариями переднего мозга. В его крыше расположены эпифиз (эндокринная железа) и теменной орган, служащий дополнительным фоторецептором: он способен воспринимать сигналы опасности (быстрое затенение) и, видимо, служит рецептором, регистрирующим сезонные изменения светового режима. Дно промежуточного мозга участвует в работе эндокринной системы в качестве нейросекреторной доли гипофиза, связанной с гипоталамической областью промежуточного мозга. Этим путем гипофиз получает информацию о состоянии внешней среды, собранную органами чувств и обработанную мозговыми центрами, что позволяет ему координировать работу всей эндокринной системы. Средствами связи служат воротные системы кровеносных сосудов, существующие во всех долях гипофиза. Они, видимо, получают и передают информацию, кодированную в химических соединениях.
Средний мозг уменьшается в размерах, теряет свое значение ведущего отдела, уменьшается его роль и как зрительного центра.
Мозжечок развит сравнительно лучше, чем у амфибий.
· Для мозга птиц характерно дальнейшее увеличение его общего объема и огромный размер переднего мозга, прикрывающего собой все остальные отделы, кроме мозжечка.
Увеличение переднего мозга , который, как и у рептилий, является ведущим отделом головного мозга, происходит за счет дна, где сильно развиваются полосатые тела. Крыша переднего мозга развита слабо, имеет небольшую толщину. Кора не получает дальнейшего развития, даже подвергается обратному развитию - исчезает латеральный участок коры.
Промежуточный мозг мал, эпифиз развит слабо, гипофиз выражен хорошо.
В среднем мозге развиты зрительные доли, т.к. зрение играет ведущую роль в жизни птиц.
Мозжечок достигает огромных размеров, имеет сложное строение. В нем различают среднюю часть и боковые выступы. Развитие мозжечка связано с полетом.
· К маммальному типу мозга относят мозг млекопитающих.
Эволюция головного мозга пошла в направлении развития крыши переднего мозга и полушарий, увеличения поверхности переднего мозга за счет извилин и борозд коры.
На всей поверхности крыши появляется слой серого вещества – настоящая кора. Это совершенно новая структура, возникающая в процессе эволюции нервной системы. У низших млекопитающих поверхность коры гладкая, а у высших - она образует многочисленные извилины, резко увеличивающие ее поверхность.
Передний мозг увеличивается в размерах за счёт крыши, приобретает значение ведущего отдела головного мозга за счет развития коры, что является характерным для маммального типа. Обонятельные доли так же сильно развиты, так как у многих млекопитающих являются органом чувств.
Кора приобретает значение ведущего отдела головного мозга, что является характерным для маммального типа мозга.
Промежуточный мозг имеет характерные придатки - эпифиз, гипофиз.
Средний мозг уменьшен в размерах. Его крыша, кроме продольной борозды, имеет еще и поперечную. Поэтому вместо двух полушарий (зрительные доли) образуется четыре бугра. Передние бугры связаны со зрительными рецепторами, а задние - со слуховыми.
Мозжечок прогрессивно развивается, что выражается в резком увеличении размеров органа и его сложной внешней и внутренней структуре.
В продолговатом мозгу по бокам обособляется путь нервных волокон, ведущих к мозжечку, а на нижней поверхности - продольные валики (пирамиды).
Пороки развития головного мозга:
Анэнцефалия (недоразвитие переднего мозга),
Микроцефалия (общее недоразвитие головного мозга),
Гидроцефалия (водянка головного мозга),
Недоразвитие лобных долей
Черепно- и спинномозговые грыжи.











