Sistema cardiovascolare dei pesci. Circoli circolatori nell'uomo: evoluzione, struttura e lavoro delle grandi e piccole caratteristiche aggiuntive Gli anfibi hanno 1 circolo circolatorio
© Utilizzo dei materiali del sito solo in accordo con l'amministrazione.
Nel corpo umano, il sistema circolatorio è progettato per soddisfare pienamente i suoi bisogni interni. Un ruolo importante nel movimento del sangue è giocato dalla presenza di un sistema chiuso in cui sono separati i flussi sanguigni arteriosi e venosi. E questo avviene grazie alla presenza di circoli circolatori.
Riferimento storico
In passato, quando gli scienziati non avevano ancora a portata di mano strumenti informativi in grado di studiare i processi fisiologici in un organismo vivente, i più grandi scienziati erano costretti a cercare caratteristiche anatomiche nei cadaveri. Naturalmente, il cuore di una persona deceduta non si contrae, quindi alcune sfumature dovevano essere capite da sole e talvolta semplicemente fantasticate. Quindi, nel II secolo d.C Claudio Galeno, autodidatta Ippocrate, presumevano che le arterie contenessero aria invece che sangue nel loro lume. Nei secoli successivi furono fatti molti tentativi per combinare e collegare insieme i dati anatomici esistenti dal punto di vista fisiologico. Tutti gli scienziati sapevano e capivano come funziona il sistema circolatorio, ma come funziona?
Gli scienziati hanno dato un enorme contributo alla sistematizzazione dei dati sulla funzione cardiaca. Miguel Servet e William Harvey nel XVI secolo. Harvey, scienziato che per primo descrisse la circolazione sistemica e polmonare , nel 1616 determinò la presenza di due cerchi, ma non riuscì a spiegare nelle sue opere come i letti arteriosi e venosi fossero collegati tra loro. E solo più tardi, nel XVII secolo, Marcello Malpighi, uno dei primi ad utilizzare il microscopio nella sua pratica, scoprì e descrisse la presenza di minuscoli capillari, invisibili ad occhio nudo, che fungono da anello di congiunzione nella circolazione sanguigna.
Filogenesi, ovvero l'evoluzione della circolazione sanguigna
A causa del fatto che, man mano che gli animali della classe dei vertebrati si evolvevano, diventavano sempre più progressivi in termini anatomici e fisiologici, richiedevano una struttura complessa del sistema cardiovascolare. Pertanto, per un movimento più rapido dell'ambiente interno liquido nel corpo di un animale vertebrato, è nata la necessità di un sistema di circolazione sanguigna chiuso. Rispetto ad altre classi del regno animale (ad esempio artropodi o vermi), nei cordati compaiono i rudimenti di un sistema vascolare chiuso. E se la lancetta, ad esempio, non ha un cuore, ma c'è un'aorta addominale e dorsale, allora nei pesci, negli anfibi (anfibi), nei rettili (rettili) appare rispettivamente un cuore a due e tre camere e in Negli uccelli e nei mammiferi appare un cuore a quattro camere, la cui particolarità è che al suo interno si concentrano due circoli di circolazione sanguigna che non si mescolano tra loro.

Pertanto, la presenza di due circoli circolatori separati negli uccelli, nei mammiferi e nell'uomo, in particolare, non è altro che l'evoluzione del sistema circolatorio, necessaria per un migliore adattamento alle condizioni ambientali.
Caratteristiche anatomiche della circolazione sanguigna
Il sistema circolatorio è un insieme di vasi sanguigni, che è un sistema chiuso per l'apporto di ossigeno e sostanze nutritive agli organi interni attraverso lo scambio di gas e sostanze nutritive, nonché per la rimozione dell'anidride carbonica e di altri prodotti metabolici dalle cellule. Il corpo umano è caratterizzato da due circoli: il circolo sistemico, o circolo grande, e il circolo polmonare, detto anche circolo piccolo.

Video: circoli di circolazione sanguigna, mini-lezione e animazione
Circolazione sistemica
La funzione principale del cerchio grande è garantire lo scambio di gas in tutti gli organi interni tranne i polmoni. Inizia nella cavità del ventricolo sinistro; rappresentato dall'aorta e dai suoi rami, dal letto arterioso del fegato, dai reni, dal cervello, dai muscoli scheletrici e da altri organi. Inoltre, questo circolo continua con la rete capillare e il letto venoso degli organi elencati; e attraverso l'ingresso della vena cava nella cavità dell'atrio destro termina in quest'ultimo.

Quindi, come già detto, l'inizio del cerchio massimo è la cavità del ventricolo sinistro. Qui viene inviato il flusso sanguigno arterioso, che contiene più ossigeno che anidride carbonica. Questo flusso entra nel ventricolo sinistro direttamente dal sistema circolatorio dei polmoni, cioè dal piccolo circolo. Il flusso arterioso proveniente dal ventricolo sinistro viene spinto attraverso la valvola aortica nel vaso più grande: l'aorta. L'aorta può essere figurativamente paragonata a una specie di albero che ha molti rami, perché da esso le arterie si estendono agli organi interni (al fegato, ai reni, al tratto gastrointestinale, al cervello - attraverso il sistema delle arterie carotidi, ai muscoli scheletrici, alla fibra grassa sottocutanea, ecc.) Le arterie degli organi, che hanno anch'esse numerose ramificazioni e portano nomi corrispondenti alla loro anatomia, trasportano l'ossigeno a ciascun organo.
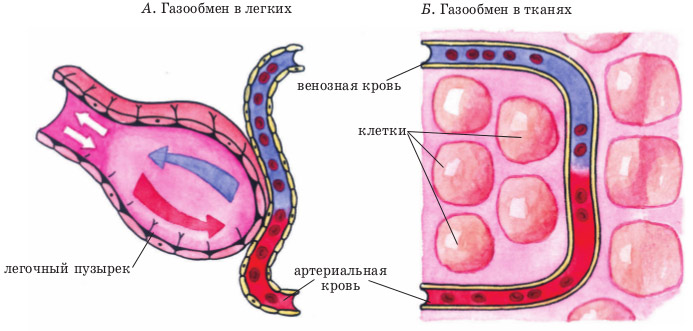
Nei tessuti degli organi interni, i vasi arteriosi sono divisi in vasi di diametro sempre più piccolo e, di conseguenza, si forma una rete capillare. I capillari sono i vasi più piccoli, praticamente senza uno strato muscolare medio, e sono rappresentati da una membrana interna - intima, rivestita da cellule endoteliali. Gli spazi tra queste cellule a livello microscopico sono così grandi rispetto ad altri vasi che consentono a proteine, gas e persino elementi formati di penetrare facilmente nel fluido intercellulare dei tessuti circostanti. Pertanto, si verifica un intenso scambio di gas e uno scambio di altre sostanze tra il capillare con il sangue arterioso e il mezzo intercellulare liquido in un particolare organo. L'ossigeno penetra dal capillare e l'anidride carbonica, come prodotto del metabolismo cellulare, entra nel capillare. Si verifica lo stadio cellulare della respirazione.
Dopo che più ossigeno è passato nei tessuti e tutta l’anidride carbonica è stata rimossa dai tessuti, il sangue diventa venoso. Tutti gli scambi di gas avvengono con ogni nuovo afflusso di sangue e durante il periodo di tempo in cui si muove lungo il capillare verso la venula, un vaso che raccoglie il sangue venoso. Cioè, ad ogni ciclo cardiaco, in una o nell'altra parte del corpo, l'ossigeno entra nei tessuti e l'anidride carbonica viene rimossa da essi.
Queste venule si uniscono in vene più grandi e si forma un letto venoso. Le vene, simili alle arterie, prendono il nome in base all'organo in cui si trovano (renale, cerebrale, ecc.). Dai grandi tronchi venosi si formano gli affluenti della vena cava superiore e inferiore, che poi confluiscono nell'atrio destro.
Caratteristiche del flusso sanguigno negli organi del circolo sistemico

Alcuni degli organi interni hanno le loro caratteristiche. Quindi, ad esempio, nel fegato non c'è solo una vena epatica, che “porta” via da esso il flusso venoso, ma anche una vena porta, che, al contrario, porta il sangue al tessuto epatico, dove avviene la depurazione del sangue. eseguito, e solo allora il sangue si raccoglie negli affluenti della vena epatica per entrare in un grande cerchio. La vena porta porta il sangue dallo stomaco e dall'intestino, quindi tutto ciò che una persona mangia o beve deve subire una sorta di “purificazione” nel fegato.
Oltre al fegato, esistono alcune sfumature in altri organi, ad esempio nei tessuti dell'ipofisi e dei reni. Così, nell'ipofisi si nota la presenza di una cosiddetta rete capillare “meravigliosa”, perché le arterie che portano il sangue all'ipofisi dall'ipotalamo sono divise in capillari, che poi si raccolgono in venule. Le venule, dopo aver raccolto il sangue con le molecole degli ormoni rilascianti, vengono nuovamente divise in capillari, e quindi si formano le vene che trasportano il sangue dalla ghiandola pituitaria. Nei reni, la rete arteriosa è divisa due volte in capillari, che sono associati ai processi di escrezione e riassorbimento nelle cellule renali - nei nefroni.
Circolazione polmonare
La sua funzione è quella di effettuare processi di scambio di gas nel tessuto polmonare al fine di saturare il sangue venoso “di scarto” con molecole di ossigeno. Inizia nella cavità del ventricolo destro, dove il flusso sanguigno venoso con una quantità estremamente piccola di ossigeno e un grande contenuto di anidride carbonica entra dalla camera atriale destra (dal “punto finale” del circolo massimo). Questo sangue si muove attraverso la valvola polmonare in uno dei grandi vasi chiamati tronco polmonare. Successivamente il flusso venoso si muove lungo il letto arterioso nel tessuto polmonare, che si divide anch'esso in una rete di capillari. Per analogia con i capillari in altri tessuti, in essi avviene lo scambio di gas, solo le molecole di ossigeno entrano nel lume del capillare e l'anidride carbonica penetra negli alveolociti (cellule degli alveoli). Ad ogni atto respiratorio, l'aria entra negli alveoli dall'ambiente, da cui l'ossigeno penetra attraverso le membrane cellulari nel plasma sanguigno. Durante l'espirazione, l'anidride carbonica che entra negli alveoli viene espulsa con l'aria espirata.

Dopo essere stato saturo di molecole di O2, il sangue acquisisce le proprietà del sangue arterioso, scorre attraverso le venule e infine raggiunge le vene polmonari. Questi ultimi, costituiti da quattro o cinque pezzi, si aprono nella cavità dell'atrio sinistro. Di conseguenza, il sangue venoso scorre attraverso la metà destra del cuore e il sangue arterioso scorre attraverso la metà sinistra; e normalmente questi flussi non dovrebbero mescolarsi.
Il tessuto polmonare ha una doppia rete di capillari. Con l'aiuto del primo, vengono effettuati processi di scambio di gas per arricchire il flusso venoso con molecole di ossigeno (relazione direttamente con il piccolo cerchio), e nel secondo, il tessuto polmonare stesso viene fornito di ossigeno e sostanze nutritive (relazione con il cerchio grande).
Cerchi di circolazione aggiuntivi
Questi concetti vengono utilizzati per distinguere l'afflusso di sangue ai singoli organi. Ad esempio, al cuore, che ha bisogno di ossigeno più di altri, l'afflusso arterioso viene effettuato dai rami dell'aorta all'inizio, che sono chiamati arterie coronarie (coronarie) destra e sinistra. Nei capillari miocardici avviene un intenso scambio di gas e il deflusso venoso avviene nelle vene coronarie. Questi ultimi si raccolgono nel seno coronarico, che sbocca direttamente nella camera atriale destra. In questo modo viene effettuato circolazione cardiaca o coronarica.

circolo coronarico (coronarico) della circolazione sanguigna nel cuore
Circolo di Willisè una rete arteriosa chiusa di arterie cerebrali. Il midollo fornisce un ulteriore apporto di sangue al cervello quando il flusso sanguigno cerebrale attraverso altre arterie viene interrotto. Questo protegge un organo così importante dalla mancanza di ossigeno o dall'ipossia. La circolazione cerebrale è rappresentata dal segmento iniziale dell'arteria cerebrale anteriore, dal segmento iniziale dell'arteria cerebrale posteriore, dalle arterie comunicanti anteriore e posteriore e dalle arterie carotidi interne.

Circolo di Willis nel cervello (variante classica della struttura)
Circolazione placentare funziona solo durante la gravidanza da parte di una donna e svolge la funzione di "respirazione" in un bambino. La placenta si forma a partire dalla 3a-6a settimana di gravidanza e inizia a funzionare pienamente a partire dalla 12a settimana. A causa del fatto che i polmoni del feto non funzionano, l'ossigeno entra nel suo sangue attraverso il flusso di sangue arterioso nella vena ombelicale del bambino.

circolazione fetale prima della nascita
Pertanto, l'intero sistema circolatorio umano può essere suddiviso in sezioni separate e interconnesse che svolgono le loro funzioni. Il corretto funzionamento di tali aree, o circoli di circolazione sanguigna, è la chiave per il sano funzionamento del cuore, dei vasi sanguigni e dell'intero corpo nel suo insieme.
Nel sistema circolatorio dei pesci, rispetto alle lancette, appare un vero cuore. Si compone di due camere, ad es. il cuore del pesce è a due camere. La prima camera è l'atrio, la seconda camera è il ventricolo del cuore. Il sangue entra prima nell'atrio, quindi viene spinto nel ventricolo dalla contrazione muscolare. Inoltre, come risultato della sua contrazione, si riversa in un grande vaso sanguigno.
Il cuore del pesce si trova nel sacco pericardico, situato dietro l'ultimo paio di archi branchiali nella cavità corporea.
Come tutti i cordati, il sistema circolatorio del pesce è chiuso. Ciò significa che in nessun punto del suo percorso il sangue lascia i vasi sanguigni e scorre nelle cavità del corpo. Per garantire lo scambio di sostanze tra il sangue e le cellule di tutto il corpo, le grandi arterie (vasi che trasportano sangue ossigenato) si ramificano gradualmente in quelle più piccole. I vasi più piccoli sono i capillari. Dopo aver ceduto l'ossigeno e assorbito l'anidride carbonica, i capillari si uniscono nuovamente in vasi più grandi (ma già venosi).
Solo nel pesce un circolo della circolazione sanguigna. Con un cuore a due camere, non può essere diversamente. Nei vertebrati più altamente organizzati (a cominciare dagli anfibi) appare una seconda circolazione (polmonare). Ma questi animali hanno anche un cuore a tre o addirittura quattro camere.
Il sangue venoso scorre attraverso il cuore, donando ossigeno alle cellule del corpo. Successivamente, il cuore spinge questo sangue nell’aorta addominale, che va alle branchie e si ramifica nelle arterie branchiali afferenti (ma nonostante il nome “arterie” contengono sangue venoso). Nelle branchie (in particolare, nei filamenti branchiali), l'anidride carbonica viene rilasciata dal sangue nell'acqua e l'ossigeno fuoriesce dall'acqua nel sangue. Ciò accade a causa della differenza nella loro concentrazione (i gas disciolti vanno dove ce ne sono meno). Arricchito di ossigeno, il sangue diventa arterioso. Le arterie branchiali efferenti (già con sangue arterioso) confluiscono in un grande vaso: l'aorta dorsale. Corre sotto la spina dorsale lungo il corpo del pesce e da esso hanno origine vasi più piccoli. Anche le arterie carotidi si diramano dall'aorta dorsale, conducendo alla testa e fornendo sangue, compreso il cervello.
Prima di entrare nel cuore, il sangue venoso passa attraverso il fegato, dove viene depurato dalle sostanze nocive.
Ci sono lievi differenze nel sistema circolatorio dei pesci ossei e cartilaginei. Ciò riguarda principalmente il cuore. Nei pesci cartilaginei (e in alcuni pesci ossei) la porzione espansa dell'aorta addominale si contrae insieme al cuore, ma nella maggior parte dei pesci ossei ciò non avviene.
Il sangue dei pesci è rosso, contiene globuli rossi con emoglobina, che lega l'ossigeno. Tuttavia, i globuli rossi dei pesci sono di forma ovale e non a forma di disco (come, ad esempio, negli esseri umani). La quantità di sangue che scorre attraverso il sistema circolatorio è inferiore nei pesci che nei vertebrati terrestri.
Il cuore del pesce non batte spesso (circa 20-30 battiti al minuto) e il numero di contrazioni dipende dalla temperatura ambiente (più è calda, più spesso). Pertanto, il loro sangue non scorre così velocemente e quindi il loro metabolismo è relativamente lento. Ciò, ad esempio, influisce sul fatto che i pesci sono animali a sangue freddo.
Nei pesci gli organi emopoietici sono la milza e il tessuto connettivo dei reni.
Nonostante il fatto che il sistema circolatorio descritto dei pesci sia caratteristico della stragrande maggioranza di essi, nei dipnoi e nei pesci con pinne lobate è leggermente diverso. Nei dipnoi, nel cuore appare un setto incompleto e appare una parvenza di (seconda) circolazione polmonare. Ma questo cerchio non passa attraverso le branchie, ma attraverso la vescica natatoria, trasformata in polmone.
Naturalmente, i pesci e gli altri abitanti acquatici hanno un cuore che ha caratteristiche simili a quelle umane, e svolge la sua funzione principale: fornire sangue al corpo. A differenza del sistema circolatorio umano, i pesci hanno un solo cerchio ed è chiuso. Nei pesci cartilaginei semplici, il flusso sanguigno avviene in linee rette, mentre nei pesci cartilaginei superiori segue la forma della lettera inglese S. Questa differenza è dovuta a una struttura più complessa e diversa. All'inizio dell'articolo vedremo il cuore dei pesci semplici, per poi passare agli straordinari abitanti cartilaginei del mondo acquatico.
Organo importante
Il cuore è l'organo principale e principale di ogni pesce, come gli esseri umani e altri animali... Può sembrare strano, perché i pesci sono animali a sangue freddo, a differenza di noi. Questo organo è una sacca muscolare che si contrae costantemente, pompando così il sangue in tutto il corpo.
Puoi scoprire che tipo di cuore ha un pesce e come si muove il sangue leggendo le informazioni in questo articolo.
Dimensioni dell'organo
La dimensione del cuore dipende dal peso corporeo totale, quindi più grande è il pesce, maggiore è il suo “motore”. Il nostro cuore è paragonato alle dimensioni di un pugno; i pesci non hanno questa opportunità. Ma come sai dalle lezioni di biologia, i piccoli pesci hanno un cuore grande solo pochi centimetri. Ma nei grandi rappresentanti del mondo sottomarino, l'organo può raggiungere anche dai venti ai trenta centimetri. Tali pesci includono pesce gatto, luccio, carpa, storione e altri.

Dov'è il cuore?
Se qualcuno è preoccupato per la domanda su quanti cuori ha un pesce, risponderemo immediatamente: uno. È sorprendente che questa domanda possa sorgere, ma come dimostra la pratica, è possibile. Molto spesso, quando puliscono il pesce, le casalinghe non sospettano nemmeno di poter trovare facilmente il cuore. Come negli esseri umani, il cuore dei pesci si trova nella parte anteriore del corpo. Per essere più precisi, proprio sotto le branchie. Il cuore è protetto su entrambi i lati da costole, proprio come le nostre. Nell'immagine che vedi sotto, l'organo principale del pesce è indicato con il numero uno.

Struttura
Considerando la respirazione dei pesci e la presenza delle branchie, il cuore è strutturato diversamente da quello degli animali terrestri. Visivamente, la forma del cuore di un pesce è simile alla nostra. Questo organo è una piccola sacca rossa, con sotto una piccola sacca rosa pallido.
Il cuore degli abitanti acquatici a sangue freddo ha solo due camere. Vale a dire il ventricolo e l'atrio. Si trovano molto vicini, o per essere più precisi, uno sopra l'altro. Il ventricolo si trova sotto l'atrio e ha una tonalità più chiara. I pesci hanno un cuore costituito da tessuto muscolare, questo è dovuto al fatto che agisce come una pompa e si contrae continuamente.
Schema di circolazione
Il cuore del pesce è collegato alle branchie tramite arterie che si trovano su entrambi i lati dell'arteria addominale principale. È chiamata anche aorta addominale; inoltre, sottili vene attraverso le quali scorre il sangue conducono da tutto il corpo all'atrio.
Il sangue del pesce è saturo di anidride carbonica, che deve essere trattato come segue. Passando attraverso le vene, il sangue entra nel cuore del pesce, dove, con l'aiuto dell'atrio, viene pompato attraverso le arterie fino alle branchie. Le branchie, a loro volta, sono dotate di numerosi capillari sottili. Questi capillari corrono attraverso le branchie e aiutano a trasportare rapidamente il sangue pompato. Successivamente, è nelle branchie che l'anidride carbonica viene miscelata e trasformata in ossigeno. Ecco perché è importante che l'acqua in cui vivono i pesci sia satura di ossigeno.
Il sangue ossigenato continua il suo viaggio attraverso il corpo del pesce e viene inviato all'aorta principale, che si trova sopra la cresta. Molti capillari si diramano da questa arteria. In essi inizia la circolazione sanguigna, o meglio lo scambio, perché, come ricordiamo, il sangue saturo di ossigeno ritorna dalle branchie.
Il risultato è la sostituzione del sangue nel corpo del pesce. Il sangue delle arterie, che di solito appare di colore rosso intenso, si trasforma in sangue delle vene, che è molto più scuro.

Direzione della circolazione sanguigna
I Pesci sono rappresentati da un atrio e un ventricolo, dotati di valvole speciali. È grazie a queste valvole che il sangue scorre in una sola direzione, escluso il flusso inverso. Questo è molto importante per un organismo vivente.
Le vene dirigono il sangue nell'atrio e da lì scorre nella seconda camera del cuore del pesce e poi verso organi speciali: le branchie. L'ultimo movimento avviene con l'aiuto dell'aorta addominale principale. Quindi puoi vedere che il cuore del pesce fa molte contrazioni infinite.

Cuore di pesce cartilagineo
Questo particolare è caratterizzato dalla presenza di cranio, spina dorsale e branchie piatte. I rappresentanti più famosi di questa classe sono gli squali e le razze.
Come i loro parenti cartilaginei, il cuore dei pesci cartilaginei ha due camere e una. Il processo di scambio di anidride carbonica con ossigeno avviene nello stesso modo descritto sopra, solo con diverse caratteristiche. Questi includono la presenza di uno spray che aiuta l'acqua a entrare nelle branchie. E tutto perché le branchie di questi pesci si trovano nella regione addominale.
Un'altra caratteristica distintiva può essere considerata la presenza di un organo come la milza. A sua volta, è la fermata finale del sangue. Ciò è necessario affinché al momento dell'attività speciale vi sia un rapido apporto di quest'ultimo all'organo desiderato.
Il sangue dei pesci cartilaginei è più saturo di ossigeno a causa dell'elevato numero di globuli rossi. E tutto a causa della maggiore attività dei reni, dove avviene la loro produzione.
Il sangue svolge numerose funzioni solo quando si muove attraverso i vasi. Lo scambio di sostanze tra il sangue e altri tessuti del corpo avviene nella rete capillare. Caratterizzato dalla sua grande lunghezza e ramificazione, offre una grande resistenza al flusso sanguigno. La pressione necessaria per superare la resistenza vascolare è creata principalmente dal cuore,
La struttura del cuore dei pesci è più semplice di quella dei vertebrati superiori. La prestazione del cuore come pompa a pressione nei pesci è significativamente inferiore rispetto agli animali terrestri. Tuttavia, affronta i suoi compiti. L'ambiente acquatico crea condizioni favorevoli per il funzionamento del cuore. Se negli animali terrestri una parte significativa del lavoro del cuore viene spesa per superare le forze di gravità e i movimenti verticali del sangue, nei pesci il denso ambiente acquatico neutralizza significativamente gli effetti gravitazionali. Un corpo allungato orizzontalmente, un piccolo volume di sangue e la presenza di un solo circuito circolatorio facilitano ulteriormente le funzioni del cuore nei pesci.
§trenta. STRUTTURA DEL CUORE
Il cuore del pesce è piccolo e rappresenta circa lo 0,1% del peso corporeo. Ci sono, ovviamente, delle eccezioni a questa regola. Ad esempio, nei pesci volanti la massa cardiaca raggiunge il 2,5% del peso corporeo.
Tutti i pesci hanno un cuore a due camere. Tuttavia, ci sono differenze tra specie nella struttura di questo organo. In termini generali, possiamo immaginare due diagrammi della struttura del cuore nella classe dei pesci. Sia nel primo che nel secondo caso si distinguono 4 cavità: il seno venoso, l'atrio, il ventricolo ed una formazione che ricorda vagamente l'arco aortico negli animali a sangue caldo - il bulbo arterioso nei teleostei e il cono arterioso negli elasmobranchi (Fig 7.1).
La differenza fondamentale tra questi schemi risiede nelle caratteristiche morfofunzionali dei ventricoli e delle formazioni arteriose.
Nei teleostei il bulbo arterioso è rappresentato da tessuto fibroso con struttura spugnosa dello strato interno, ma privo di valvole.
Negli elasmobranchi, il cono arterioso, oltre al tessuto fibroso, contiene anche il tipico tessuto muscolare cardiaco e quindi è dotato di contrattilità. Il cono ha un sistema di valvole che facilitano il movimento unidirezionale del sangue attraverso il cuore.
Riso. 7.1. Schema della struttura del cuore del pesce
Differenze nella struttura del miocardio sono state riscontrate nel ventricolo del cuore di pesce. È generalmente accettato che il miocardio dei pesci sia specifico e sia rappresentato da tessuto cardiaco omogeneo, uniformemente penetrato da trabecole e capillari. Il diametro delle fibre muscolari nei pesci è inferiore a quello degli animali a sangue caldo ed è di 6-7 micron, ovvero la metà, ad esempio, del miocardio di un cane. Tale miocardio è chiamato spugnoso.
I resoconti sulla vascolarizzazione miocardica dei pesci sono piuttosto confusi. Il miocardio è rifornito di sangue venoso dalle cavità trabecolari, che a loro volta si riempiono di sangue proveniente dal ventricolo attraverso i vasi tebesi. Nel senso classico, i pesci non hanno una circolazione coronarica. Almeno, i cardiologi aderiscono a questo punto di vista. Tuttavia, nella letteratura sull'ittiologia si trova spesso il termine “circolazione coronarica dei pesci”.
Negli ultimi anni, i ricercatori hanno scoperto molte variazioni nella vascolarizzazione del miocardio. Ad esempio, S. Agnisola et. al (1994) hanno riportato la presenza di un miocardio a doppio strato nella trota e nella razza elettrica. Sul lato endocardico si trova uno strato spugnoso e sopra di esso uno strato di fibre miocardiche con disposizione compatta e ordinata.
Gli studi hanno dimostrato che lo strato spugnoso del miocardio viene rifornito di sangue venoso dalle lacune trabecolari e lo strato compatto riceve sangue arterioso attraverso le arterie ipobronchiali del secondo paio di pustole branchiali. Negli elasmobranchi, la circolazione coronarica differisce in quanto il sangue arterioso dalle arterie ipobronchiali raggiunge lo strato spugnoso attraverso un sistema capillare ben sviluppato ed entra nella cavità ventricolare attraverso i vasi di Tibesio.
Un'altra differenza significativa tra teleostei ed elasmobranchi è la morfologia del pericardio.
Nei teleostei il pericardio somiglia a quello degli animali terrestri. È rappresentato da un guscio sottile.
Negli elasmobranchi il pericardio è formato da tessuto cartilagineo, quindi è come una capsula dura ma elastica. In quest'ultimo caso, durante la diastole, si crea un certo vuoto nello spazio pericardico, che facilita l'afflusso di sangue al seno venoso e all'atrio senza ulteriore dispendio energetico.
§31. PROPRIETÀ ELETTRICHE DEL CUORE
La struttura dei miociti del muscolo cardiaco dei pesci è simile a quella dei vertebrati superiori. Pertanto, le proprietà elettriche del cuore sono simili. Il potenziale di riposo dei miociti nei teleostei e negli elasmobranchi è di 70 mV e nelle missine è di 50 mV. Al picco del potenziale d'azione, viene registrata una variazione nel segno e nell'entità del potenziale da meno 50 mV a più 15 mV. La depolarizzazione della membrana del miocita porta all'eccitazione dei canali sodio-calcio. Innanzitutto, gli ioni sodio e poi gli ioni calcio si riversano nella cellula del miocita. Questo processo è accompagnato dalla formazione di un plateau allungato e la refrattarietà assoluta del muscolo cardiaco viene registrata funzionalmente. Questa fase nei pesci è molto più lunga: circa 0,15 s.
La successiva attivazione dei canali del potassio e il rilascio di ioni potassio dalla cellula assicurano una rapida ripolarizzazione della membrana del miocita. A sua volta, la ripolarizzazione della membrana chiude i canali del potassio e apre i canali del sodio. Di conseguenza, il potenziale della membrana cellulare ritorna al livello originale di meno 50 mV.
I miociti del cuore del pesce, capaci di generare potenziale, sono localizzati in alcune aree del cuore, che sono collettivamente combinate nel “sistema di conduzione del cuore”. Come nei vertebrati superiori, nei pesci l'inizio della sistole cardiaca avviene nel nodo sinatriale.
A differenza degli altri vertebrati, nei pesci il ruolo di pacemaker è svolto da tutte le strutture del sistema di conduzione, che nei teleostei comprende il centro del condotto uditivo, un nodo del setto atrioventricolare, da cui si estendono le cellule di Purkinje ai tipici cardiociti del ventricolo .
La velocità di eccitazione attraverso il sistema di conduzione del cuore nei pesci è inferiore a quella dei mammiferi e varia nelle diverse parti del cuore. La massima velocità di propagazione potenziale è stata registrata nelle strutture del ventricolo.
L'elettrocardiogramma del pesce assomiglia all'elettrocardiogramma umano nelle derivazioni V3 e V4 (Fig. 7.2). Tuttavia, la tecnica di applicazione del piombo per i pesci non è stata sviluppata in modo così dettagliato come per i vertebrati terrestri.

Riso. 7.2. Elettrocardiogramma dei pesci
Nella trota e nell'anguilla, sull'elettrocardiogramma sono ben visibili le onde P, Q, R, S e T. Solo l'onda S appare ipertrofica, e l'onda Q ha inaspettatamente direzione positiva; negli elasmobranchi, oltre ai classici cinque denti , l'elettrocardiogramma rivela le onde Bd tra i denti S e T, così come il dente Br tra i denti G e R. Nell'elettrocardiogramma di un'anguilla, l'onda P è preceduta da un'onda V. L'eziologia delle onde è la seguente:
l'onda P corrisponde all'eccitazione del condotto uditivo e alla contrazione del seno venoso e dell'atrio;
il complesso QRS caratterizza l'eccitazione del nodo atrioventricolare e della sistole ventricolare;
L'onda T si verifica in risposta alla ripolarizzazione delle membrane cellulari del ventricolo cardiaco.
§32. LAVORO DEL CUORE
Il cuore del pesce funziona ritmicamente. La frequenza cardiaca dei pesci dipende da molti fattori.
Frequenza cardiaca (battiti al minuto) nella carpa a 20 °C
Larva
Giovani del peso di 0,02 g 80
Piccoli avannotti del peso di 25 g 40
Bambini di due anni di peso 500 g 30
Negli esperimenti in vitro (cuore isolato e perfuso), la frequenza cardiaca della trota iridea e della razza elettrica era di 20-40 battiti al minuto.
Tra i tanti fattori, la temperatura dell’ambiente ha l’effetto più pronunciato sulla frequenza cardiaca. Utilizzando il metodo telemetrico su spigole e passere, è stata rilevata la seguente relazione (Tabella 7.1).
7.1. Dipendenza della frequenza cardiaca dalla temperatura dell'acqua
| Temperatura, °C | Frequenza cardiaca, battiti al minuto | Temperatura, °C | Frequenza cardiaca, battiti al minuto |
| 11,5 | |||
È stata stabilita la sensibilità delle specie dei pesci ai cambiamenti di temperatura. Pertanto, nella passera, quando la temperatura dell'acqua aumenta da g a 12 °C, la frequenza cardiaca aumenta di 2 volte (da 24 a 50 battiti al minuto), nel pesce persico - solo da 30 a 36 battiti al minuto.
La regolazione delle contrazioni cardiache viene effettuata utilizzando il sistema nervoso centrale e i meccanismi intracardiaci. Come negli animali a sangue caldo, la tachicardia è stata osservata nei pesci in esperimenti in vivo quando la temperatura del sangue che scorreva al cuore aumentava. Una diminuzione della temperatura del sangue che scorre al cuore ha causato bradicardia. La vagotomia ha ridotto il livello di tachicardia.
Molti fattori umorali hanno anche un effetto cronotropo. Un effetto cronotropo positivo è stato ottenuto con la somministrazione di atropina, adrenalina ed eptatretina. La cronotropia negativa è stata causata da acetilcolina, efedrina e cocaina.
È interessante notare che lo stesso agente umorale a diverse temperature ambientali può avere esattamente l'effetto opposto sul cuore dei pesci. Pertanto, su un cuore di trota isolato a bassa temperatura (6°C), l'adrenalina provoca un effetto cronotropo positivo e, sullo sfondo delle temperature elevate (15°C) del fluido di perfusione, un effetto cronotropo negativo.
La gittata cardiaca cardiaca nei pesci è stimata in 15-30 ml/kg al minuto. La velocità lineare del sangue nell'aorta addominale è di 8-20 cm/s. In vitro sulla trota è stata stabilita la dipendenza della gittata cardiaca dalla pressione del fluido di perfusione e dal contenuto di ossigeno in esso contenuto. Tuttavia, nelle stesse condizioni, il volume minuto della pastinaca elettrica non è cambiato.
I ricercatori includono più di una dozzina di componenti nel perfusato.
Composizione del perfusato per cuore di trota (g/l)
Cloruro di sodio 7,25
Cloruro di potassio 0,23
Fluoruro di calcio 0,23
Solfato di magnesio (cristallino) 0,23
Sodio fosfato monosostituito (cristallino) 0,016
Fosfato disodico (cristallino) 0,41
Glucosio 1.0
Idolo di polivinilpirrolo (PVP) colloidale 10.0
Appunti:
I. La soluzione è saturata con una miscela di gas composta da 99,5% di ossigeno, 0,5% di anidride carbonica (anidride carbonica) o una miscela di aria (99,5%) con anidride carbonica (0,5%).
2. Il pH del perfusato viene regolato a 7,9 a una temperatura di 10 °C utilizzando bicarbonato di sodio.
Composizione del perfusato per cuore di razza elettrica (g/l)
Cloruro di sodio 16.36
Cloruro di potassio 0,45
Cloruro di magnesio 0,61
Solfato di sodio 0,071
Sodio fosfato monosostituito (cristallino) 0,14
Bicarbonato di sodio 0,64
Urea 21,0
Glucosio 0,9
Appunti:
1. Il perfusato è saturo della stessa miscela di gas. 2.pH 7,6.
In tali soluzioni, il cuore di pesce isolato conserva le sue proprietà e funzioni fisiologiche per un tempo molto lungo. Quando si eseguono semplici manipolazioni con il cuore, è consentito l'uso della soluzione isotonica di cloruro di sodio. Tuttavia, non dovresti contare sul lavoro prolungato del muscolo cardiaco.
§33. CERCHIO DELLA CIRCOLAZIONE DEL SANGUE
I pesci, come sai, hanno un circolo di circolazione sanguigna. Eppure il sangue circola più a lungo. Una circolazione sanguigna completa nei pesci dura circa 2 minuti (nell'uomo il sangue passa attraverso due circoli di circolazione sanguigna in 20-30 secondi). Dal ventricolo, attraverso il bulbo arterioso o il cono arterioso, il sangue entra nella cosiddetta aorta addominale, che si estende dal cuore in direzione craniale fino alle branchie (Fig. 7.3).
L'aorta addominale è divisa in arterie branchiali afferenti sinistra e destra (a seconda del numero di archi branchiali). Da loro un'arteria petalo si estende a ciascun filamento branchiale, e da esso a ciascun petalo partono due arteriole, che formano una rete capillare dei vasi più fini, la cui parete è formata da un epitelio a strato singolo con ampi spazi intercellulari. I capillari si fondono in un'unica arteriola efferente (a seconda del numero di petali). Le arteriole efferenti formano l'arteria efferente petalo. Le arterie petalo formano le arterie branchiali efferenti sinistra e destra, attraverso le quali scorre il sangue arterioso.

Riso. 7.3. Diagramma della circolazione sanguigna dei pesci ossei:
1- aorta addominale; 2 - arterie carotidi; 3 - arterie branchiali; 4- arteria e vena succlavia; b- aorta dorsale; 7 - vena cardinale posteriore; 8- vasi renali; 9 - vena della coda; 10 - vena reversibile dei reni; 11 - vasi intestinali, 12 - vena porta; 13 - vasi epatici; 14- vene epatiche; 15- venoso 16- condotto di Cuvier; 17- vena cardinale anteriore
Le arterie carotidi si estendono dalle arterie branchiali efferenti alla testa. Successivamente, le arterie branchiali si fondono per formare un unico grande vaso: l'aorta dorsale, che si estende in tutto il corpo sotto la colonna vertebrale e fornisce la circolazione sistemica arteriosa. Le principali arterie di partenza sono la succlavia, la mesenterica, iliaca, la caudale e la segmentale.
La parte venosa del circolo inizia con i capillari dei muscoli e degli organi interni, che si uniscono per formare vene cardinali accoppiate anteriori e posteriori accoppiate. Le vene cardinali si uniscono alle due vene epatiche per formare i dotti di Cuvier, che sfociano nel seno venoso.
Pertanto, il cuore dei pesci pompa e succhia solo sangue venoso. Tuttavia, tutti gli organi e i tessuti ricevono sangue arterioso, poiché prima di riempire la microvascolarizzazione degli organi, il sangue passa attraverso l'apparato branchiale, nel quale avvengono gli scambi di gas tra il sangue venoso e l'ambiente acquatico.
§34. MOVIMENTO DEL SANGUE E PRESSIONE SANGUIGNA
Il sangue si muove attraverso i vasi a causa della differenza di pressione all'inizio e alla fine della circolazione sanguigna. Quando la pressione sanguigna è stata misurata senza anestesia in posizione ventrale (causando bradicardia), il salmone nell'aorta addominale era 82/50 mm Hg. Art., e nella dorsale 44/37 mm Hg. Arte. Uno studio su pesci anestetizzati di diverse specie ha dimostrato che l'anestesia riduce significativamente la pressione sistolica, fino a 30-70 mmHg. Arte. La pressione del polso varia da 10 a 30 mm Hg a seconda della specie di pesce. Arte. L'ipossia ha portato ad un aumento della pressione del polso a 40 mmHg. Arte.
Alla fine della circolazione sanguigna, la pressione sanguigna sulle pareti dei vasi sanguigni (nei dotti di Cuvier) non superava i 10 mm Hg. Arte.
La maggiore resistenza al flusso sanguigno è fornita dal sistema branchiale con i suoi capillari lunghi e altamente ramificati. Nella carpa e nella trota la differenza di pressione sistolica nell'aorta addominale e dorsale, cioè all'ingresso e all'uscita dall'apparato branchiale, è del 40-50%. Durante l'ipossia, le branchie offrono una resistenza ancora maggiore al flusso sanguigno.
Oltre al cuore, anche altri meccanismi contribuiscono al movimento del sangue attraverso i vasi. Pertanto, l’aorta dorsale, che ha la forma di un tubo diritto con pareti relativamente rigide (rispetto all’aorta addominale), offre poca resistenza al flusso sanguigno. Le arterie segmentali, caudali e le altre hanno un sistema di valvole a tasca simili a quelle dei grandi vasi venosi. Questo sistema di valvole impedisce al sangue di rifluire. Per il flusso sanguigno venoso sono di grande importanza anche le contrazioni adiacenti alle vene del topo, che spingono il sangue in direzione cardiaca.
Il ritorno venoso e la gittata cardiaca sono ottimizzati mediante la mobilitazione del sangue immagazzinato. È stato sperimentalmente dimostrato che nella trota il carico muscolare porta ad una diminuzione del volume della milza e del fegato.
Infine, il movimento del sangue è facilitato dal meccanismo di riempimento uniforme del cuore e dall'assenza di forti fluttuazioni sistolico-diastoliche della gittata cardiaca. Il riempimento del cuore è assicurato già durante la diastole ventricolare, quando si crea un certo vuoto nella cavità pericardica e il sangue riempie passivamente il seno venoso e l'atrio. Lo shock sistolico è smorzato dal bulbo arterioso, che ha una superficie interna elastica e porosa.
Capitolo 8. SCAMBIO DI GAS DEI PESCE
La concentrazione di ossigeno nel serbatoio è l'indicatore più instabile dell'habitat dei pesci, che cambia molte volte durante il giorno. Tuttavia, la pressione parziale dell'ossigeno e dell'anidride carbonica nel sangue dei pesci è abbastanza stabile e appartiene alle rigide costanti dell'omeostasi.
Come mezzo respiratorio, l'acqua è inferiore all'aria (Tabella 8.1).
8.1. Confronto tra acqua e aria come mezzo respiratorio (a una temperatura di 20 °C)
Date le condizioni iniziali così sfavorevoli per lo scambio di gas, l'evoluzione ha intrapreso la strada della creazione di ulteriori meccanismi di scambio di gas negli animali acquatici, che consentono loro di tollerare pericolose fluttuazioni della concentrazione di ossigeno nel loro ambiente. Oltre alle branchie, allo scambio gassoso partecipano anche la pelle, il tratto gastrointestinale, la vescica natatoria e organi speciali.
§35. GILLS - UN ORGANO EFFICACE PER LO SCAMBIO DI GAS NELL'AMBIENTE ACQUATICO
L'onere principale di fornire ossigeno al pesce e rimuovere l'anidride carbonica da esso ricade sulle branchie. Eseguono un lavoro tetanico. Se si confronta la respirazione branchiale con quella polmonare si arriva alla conclusione che i pesci devono pompare attraverso le branchie un mezzo respiratorio 30 volte maggiore in volume e 20.000 (!) volte maggiore in massa.
Un esame più attento mostra che le branchie sono ben adattate allo scambio di gas nell'ambiente acquatico. L'ossigeno passa nel letto capillare delle branchie lungo un gradiente di pressione parziale, che nei pesci è di 40-100 mm Hg. Arte. Questo è anche il motivo del passaggio dell'ossigeno dal sangue al fluido intercellulare nei tessuti.
Qui il gradiente di pressione parziale dell'ossigeno è stimato a 1–15 mmHg. Art., gradiente di concentrazione di anidride carbonica - 3-15 mmHg.
Lo scambio di gas in altri organi, ad esempio attraverso la pelle, avviene secondo le stesse leggi fisiche, ma l'intensità della diffusione in essi è molto inferiore. La superficie branchiale è 10-60 volte più grande della superficie corporea del pesce. Inoltre le branchie, organi altamente specializzati per lo scambio gassoso, avranno grandi vantaggi anche a parità di area degli altri organi.
La struttura più perfetta dell'apparato branchiale è caratteristica dei pesci ossei. La base dell'apparato branchiale sono 4 paia di archi branchiali. Sugli archi branchiali sono presenti filamenti branchiali ben vascolarizzati che formano la superficie respiratoria (Fig. 8.1).
Sul lato dell'arco branchiale rivolto verso la cavità orale si trovano strutture più piccole: i rastrelli branchiali, che sono in gran parte responsabili della purificazione meccanica dell'acqua mentre scorre dalla cavità orale ai filamenti branchiali.
I filamenti branchiali microscopici si trovano trasversalmente ai filamenti branchiali, che sono gli elementi strutturali delle branchie come organi respiratori (vedi Fig. 8.1; 8.2). L'epitelio che ricopre i petali ha tre tipi di cellule: respiratorie, mucose e di sostegno. L'area delle lamelle secondarie e, di conseguenza, dell'epitelio respiratorio dipende dalle caratteristiche biologiche del pesce: stile di vita, metabolismo basale, richiesta di ossigeno. Quindi, nel tonno con una massa di 100 g, la superficie branchiale è di 20-30 cm 2 / g, nella triglia - 10 cm 2 / g, nella trota - 2 cm 2 / g, nello scarafaggio - 1 cm 2 / g .
Lo scambio di gas branchiale può essere efficace solo con un flusso costante di acqua attraverso l'apparato branchiale. L'acqua irriga costantemente i filamenti branchiali e ciò è facilitato dall'apparato orale. L'acqua scorre dalla bocca alle branchie. La maggior parte delle specie ittiche possiede questo meccanismo.

Riso. 8.1. Struttura delle branchie dei pesci ossei:
1- filamenti branchiali; 2- filamenti branchiali; Arteria 3-branchiale; 4 - vena branchiale; arteria a 5 petali; 6 - vena del petalo; Rastrellieri a 7 branchie; Arco a 8 branchie
Tuttavia, è noto che le specie grandi e attive, come il tonno, non chiudono la bocca e non hanno movimenti respiratori delle coperture branchiali. Questo tipo di ventilazione branchiale è detta “ariete”; è possibile solo ad alte velocità di movimento nell'acqua.
Il passaggio dell'acqua attraverso le branchie e il movimento del sangue attraverso i vasi dell'apparato branchiale è caratterizzato da un meccanismo controcorrente, che garantisce un'altissima efficienza di scambio di gas. Dopo aver attraversato le branchie, l'acqua perde fino al 90% dell'ossigeno in essa disciolto (Tabella 8.2).
8.2. Efficienza dell'estrazione dell'ossigeno dall'acqua mediante diverse forchette da pesce,%
I filamenti e i petali branchiali si trovano molto vicini, ma a causa della bassa velocità dell'acqua che si muove attraverso di essi, non creano molta resistenza al flusso dell'acqua. Secondo i calcoli, nonostante la grande quantità di lavoro necessaria per spostare l'acqua attraverso l'apparato branchiale (almeno 1 m 3 di acqua per 1 kg di peso vivo al giorno), i costi energetici dei pesci sono bassi.
L'iniezione d'acqua è fornita da due pompe: orale e branchiale. In diverse specie di pesci, una di queste può predominare. Ad esempio, nel cefalo e nel sugarello che si muovono velocemente funziona principalmente la pompa orale, mentre nei pesci di fondo che si muovono lentamente (passera o pesce gatto) funziona la pompa branchiale.
La frequenza dei movimenti respiratori nei pesci dipende da molti fattori, ma due hanno la maggiore influenza su questo indicatore fisiologico: la temperatura dell'acqua e il contenuto di ossigeno in essa contenuto. La dipendenza della frequenza respiratoria dalla temperatura è mostrata in Fig. 8.2.
Pertanto, la respirazione branchiale dovrebbe essere considerata un meccanismo molto efficace di scambio di gas nell'ambiente acquatico in termini di efficienza dell'estrazione dell'ossigeno, nonché del consumo di energia per questo processo. Nel caso in cui il meccanismo branchiale non possa far fronte al compito di un adeguato scambio di gas, vengono attivati altri meccanismi (ausiliari). 
Riso. 8.2. Dipendenza della frequenza respiratoria dalla temperatura dell'acqua negli avannotti di carpa
§36. RESPIRAZIONE DELLA PELLE
La respirazione cutanea è sviluppata a vari livelli in tutti gli animali, ma in alcune specie di pesci potrebbe rappresentare il principale meccanismo di scambio gassoso.
La respirazione cutanea è essenziale per le specie che conducono uno stile di vita sedentario in condizioni di scarso ossigeno o che abbandonano il bacino per brevi periodi (anguilla, saltafango, pesce gatto). Nell'anguilla adulta la respirazione cutanea diventa quella principale e raggiunge il 60% del volume totale di scambio gassoso
8.3. Proporzione della respirazione cutanea in diverse specie di pesci
Lo studio dello sviluppo ontogenetico dei pesci indica che la respirazione cutanea è primaria rispetto alla respirazione branchiale. Embrioni e larve di pesci scambiano gas con l'ambiente attraverso il tessuto tegumentario. L'intensità della respirazione cutanea aumenta con l'aumentare della temperatura dell'acqua, poiché l'aumento della temperatura aumenta il metabolismo e riduce la solubilità dell'ossigeno nell'acqua.
In generale, l'intensità dello scambio gassoso cutaneo è determinata dalla morfologia della pelle. Nelle anguille, la pelle presenta vascolarizzazione e innervazione ipertrofica rispetto ad altre specie.
In altre specie, come gli squali, la percentuale di respirazione cutanea è insignificante, ma anche la loro pelle ha una struttura ruvida con un sistema di afflusso sanguigno poco sviluppato.
L'area dei vasi sanguigni cutanei in diverse specie di pesci ossei varia da 0,5 a 1,5 cm:/g di peso vivo. Il rapporto tra l'area dei capillari cutanei e quelli branchiali varia ampiamente: da 3:1 nel cobite a 10:1 nella carpa.
Lo spessore dell'epidermide, che varia da 31-38 µm nella passera, a 263 µm nell'anguilla e 338 µm nel loach, è determinato dal numero e dalle dimensioni delle cellule della mucosa. Tuttavia, ci sono pesci con scambi di gas molto intensi sullo sfondo della normale macro e microstruttura della pelle.
In conclusione, va sottolineato che il meccanismo della respirazione cutanea negli animali evidentemente non è stato sufficientemente studiato. Un ruolo importante in questo processo è svolto dal muco cutaneo, che contiene sia l'emoglobina che l'enzima anidrasi carbonica.
§37. RESPIRAZIONE INTESTINALE
In condizioni estreme (ipossia), la respirazione intestinale viene utilizzata da molte specie di pesci. Tuttavia, ci sono pesci in cui il tratto gastrointestinale ha subito cambiamenti morfologici ai fini di un efficiente scambio di gas. In questo caso, di regola, aumenta la lunghezza dell'intestino. In tali pesci (pesce gatto, ghiozzo), l'aria viene inghiottita e diretta dai movimenti peristaltici dell'intestino verso una sezione specializzata. In questa parte del tratto gastrointestinale, la parete intestinale è adattata allo scambio di gas, in primo luogo, a causa della vascolarizzazione capillare ipertrofica e, in secondo luogo, per la presenza dell'epitelio colonnare respiratorio. Una bolla d'aria atmosferica ingerita nell'intestino si trova sotto una certa pressione, che aumenta il coefficiente di diffusione dell'ossigeno nel sangue. In questo luogo l'intestino viene rifornito di sangue venoso, quindi c'è una buona differenza nella pressione parziale di ossigeno e anidride carbonica e nell'unidirezionalità della loro diffusione. La respirazione intestinale è diffusa nel pesce gatto americano. Tra questi ci sono specie con uno stomaco adatto allo scambio di gas.
La vescica natatoria non solo fornisce al pesce una galleggiabilità neutra, ma svolge anche un ruolo nello scambio di gas. Può essere aperto (salmone) o chiuso (carpa). La vescica aperta è collegata tramite un condotto d'aria all'esofago e la sua composizione gassosa può essere rapidamente aggiornata. In una vescica chiusa, i cambiamenti nella composizione del gas avvengono solo attraverso il sangue.
.Nella parete della vescica natatoria è presente uno speciale sistema capillare, comunemente chiamato “ghiandola del gas”. I capillari della ghiandola formano anelli controcorrente fortemente curvi. L'endotelio della ghiandola gassosa è in grado di secernere acido lattico e quindi di modificare localmente il pH del sangue. Questo, a sua volta, fa sì che l’emoglobina rilasci ossigeno direttamente nel plasma sanguigno. Si scopre che il sangue che scorre dalla vescica natatoria è saturo di ossigeno. Tuttavia, il meccanismo controcorrente del flusso sanguigno nella ghiandola del gas fa sì che l'ossigeno plasmatico si diffonda nella cavità vescicale. Pertanto, la bolla crea una riserva di ossigeno, che viene utilizzata dal corpo del pesce in condizioni sfavorevoli.
Altri dispositivi per lo scambio gassoso sono rappresentati dal labirinto (gourami, lalius, galletto), organo epibranchiale (anguilla del riso), polmoni (pesce polmone), apparato orale (persico rampicante), cavità faringee (Ophiocephalus sp.). Il principio dello scambio di gas in questi organi è lo stesso dell'intestino o della vescica natatoria. La base morfologica dello scambio di gas in essi è un sistema di circolazione capillare modificato più un assottigliamento delle mucose (Fig. 8.3). 
Riso. 8.3. Tipi di organi epibranchiali:
1- trespolo cursore: 2- mucchio; 3- testa di serpente; 4-Nilo Sharmuth
Morfologicamente e funzionalmente, gli pseudobranchi - formazioni speciali dell'apparato branchiale - sono associati agli organi respiratori. Il loro ruolo non è completamente compreso. Quello. il fatto che il sangue ossigenato fluisca verso queste strutture dalle branchie lo indica. che non partecipano allo scambio di ossigeno. Tuttavia, la presenza di una grande quantità di anidrasi carbonica sulle membrane degli pseudobranchi suggerisce la partecipazione di queste strutture nella regolazione dello scambio di anidride carbonica all'interno dell'apparato branchiale.
Funzionalmente collegata agli pseudobranchi è la cosiddetta ghiandola vascolare, situata sulla parete posteriore del bulbo oculare e che circonda il nervo ottico. La ghiandola vascolare ha una rete di capillari che ricorda quella della ghiandola gassosa della vescica natatoria. C'è un punto di vista secondo cui la ghiandola vascolare garantisce l'apporto di sangue altamente ossigenato alla retina dell'occhio con il minor apporto possibile di anidride carbonica al suo interno. È probabile che la fotoricezione influenzi il pH delle soluzioni in cui si verifica. Pertanto, il sistema pseudobranchiale-ghiandola vascolare può essere considerato come un ulteriore filtro tampone della retina. Se si tiene conto che la presenza di questo sistema non è legata alla posizione tassonomica dei pesci, ma piuttosto all'habitat (questi organi sono più spesso presenti nelle specie marine che vivono in acque ad elevata trasparenza, e per le quali la visione è il canale più importante di comunicazione con l'ambiente esterno), allora questa ipotesi sembra convincente.
§38. TRASFERIMENTO DEI GAS VIA SANGUE
Non ci sono differenze fondamentali nel trasporto dei gas attraverso il sangue nei pesci. Come negli animali polmonari, nei pesci le funzioni di trasporto del sangue sono realizzate grazie all'elevata affinità dell'emoglobina per l'ossigeno, alla solubilità relativamente elevata dei gas nel plasma sanguigno e alla trasformazione chimica dell'anidride carbonica in carbonati e bicarbonati.
Il principale trasportatore di ossigeno nel sangue dei pesci è l'emoglobina. È interessante notare che l'emoglobina dei pesci è funzionalmente divisa in due tipi: acido-sensibile e acido-insensibile.
L’emoglobina acido-sensibile perde la sua capacità di legare l’ossigeno quando il pH del sangue diminuisce.
L'emoglobina, che è insensibile agli acidi, non risponde ai valori del pH e per i pesci la sua presenza è di vitale importanza, poiché la loro attività muscolare è accompagnata da grandi emissioni di acido lattico nel sangue (risultato naturale della glicolisi in condizioni di costante ipossia).
Alcune specie di pesci artici e antartici non hanno affatto emoglobina nel sangue. In letteratura sono riportati casi dello stesso fenomeno nella carpa. Esperimenti sulle trote hanno dimostrato che i pesci non soffrono di asfissia senza emoglobina funzionale (tutta l'emoglobina è stata legata artificialmente utilizzando CO) a temperature dell'acqua inferiori a 5 °C. Ciò indica che il fabbisogno di ossigeno dei pesci è significativamente inferiore a quello degli animali terrestri (specialmente a basse temperature dell'acqua, quando aumenta la solubilità dei gas nel plasma sanguigno).
In determinate condizioni, il plasma da solo è in grado di far fronte al trasporto dei gas. Tuttavia, in condizioni normali, nella stragrande maggioranza dei pesci, lo scambio di gas senza emoglobina è praticamente escluso. La diffusione dell'ossigeno dall'acqua nel sangue avviene lungo un gradiente di concentrazione. Il gradiente viene mantenuto quando l'ossigeno disciolto nel plasma è legato all'emoglobina, cioè la diffusione dell'ossigeno dall'acqua avviene fino a quando l'emoglobina è completamente satura di ossigeno. La capacità di ossigeno del sangue varia da 65 mg/l nella razza a 180 mg/l nel salmone. Tuttavia, la saturazione del sangue con anidride carbonica (anidride carbonica) può ridurre di 2 volte la capacità di ossigeno del sangue dei pesci. 
Riso. 8.4. Il ruolo dell'anidrasi carbonica nel trasporto dell'anidride carbonica nel sangue
L'anidride carbonica viene trasportata nel sangue in modo diverso. Il ruolo dell'emoglobina nel trasferimento dell'anidride carbonica sotto forma di carboemoglobina è piccolo. I calcoli mostrano che l'emoglobina trasporta non più del 15% dell'anidride carbonica generata dal metabolismo dei pesci. Il principale sistema di trasporto per il trasferimento dell'anidride carbonica è il plasma sanguigno.
Entrando nel sangue a seguito della diffusione dalle cellule, l'anidride carbonica, a causa della sua limitata solubilità, crea un aumento della pressione parziale nel plasma e quindi dovrebbe inibire la transizione del gas dalle cellule al flusso sanguigno. In realtà questo non avviene. Nel plasma, sotto l'influenza dell'anidrasi carbonica degli eritrociti, avviene la reazione
CO2 + H2O>H2 CO3 >H + + HCO3
A causa di ciò, la pressione parziale dell'anidride carbonica sulla membrana cellulare dal plasma sanguigno viene costantemente ridotta e la diffusione dell'anidride carbonica nel sangue avviene in modo uniforme. Il ruolo dell'anidrasi carbonica è mostrato schematicamente in Fig. 8.4.
Il bicarbonato risultante entra nell'epitelio branchiale con il sangue, che contiene anche anidrasi carbonica. Pertanto, i bicarbonati vengono convertiti in anidride carbonica e acqua nelle branchie. Inoltre, lungo il gradiente di concentrazione, la CO2 si diffonde dal sangue nell'acqua che lava le branchie.
L'acqua che scorre attraverso i filamenti branchiali entra in contatto con l'epitelio branchiale per non più di 1 s, quindi il gradiente di concentrazione dell'anidride carbonica non cambia e lascia il flusso sanguigno a velocità costante. L'anidride carbonica viene rimossa più o meno allo stesso modo in altri organi respiratori. Inoltre, quantità significative di anidride carbonica prodotta a seguito del metabolismo vengono escrete dal corpo sotto forma di carbonati nelle urine, nel succo pancreatico, nella bile e attraverso la pelle.
La superclasse Pesci appartiene al phylum Chordata. Vivono nell'acqua. E hanno una serie di caratteristiche associate alla vita in esso.
Sistema circolatorio dei pesci
Come tutti i cordati, i pesci hanno un sistema circolatorio chiuso. Sia nei pesci ossei che in quelli cartilaginei, il sangue proveniente dal cuore entra nei vasi sanguigni e da essi ritorna al cuore. Questi animali hanno due camere nel cuore: l'atrio e il ventricolo. Esistono tre tipi di navi:
- arterie;
- vene;
- capillari.
Le arterie trasportano il sangue lontano dal cuore e le pareti di questi vasi sono più spesse in modo da poter resistere alla pressione creata dal cuore. Attraverso le vene, il sangue ritorna al cuore e la pressione al loro interno diminuisce, quindi le loro pareti sono più sottili. E i capillari sono i vasi più piccoli, le cui pareti sono costituite da un unico strato di cellule, perché la loro funzione principale è lo scambio di gas.
Circolazione sanguigna dei pesci
Prima di considerare il processo stesso di circolazione del sangue, è necessario ricordare i tipi di sangue. Può essere arterioso, che contiene molto ossigeno, e venoso, saturo di anidride carbonica. Pertanto, il tipo di sangue non ha nulla a che fare con il nome dei vasi attraverso i quali scorre, ma solo con la sua composizione. Quanto ai pesci, hanno sangue venoso in entrambe le camere del cuore e una sola circolazione.
Consideriamo in sequenza il movimento del sangue:
- Il ventricolo si contrae e spinge il sangue venoso nelle arterie branchiali.
- Nelle branchie le arterie si diramano nei capillari. Qui avviene lo scambio gassoso e il sangue passa da venoso ad arterioso.
- Dai capillari, il sangue arterioso si raccoglie nell'aorta addominale.
- L'aorta si ramifica nelle arterie degli organi.
- Negli organi, le arterie si ramificano nuovamente nei capillari, dove il sangue, cedendo ossigeno e sottraendo anidride carbonica, diventa venoso dall'arteria.
- Il sangue venoso proveniente dagli organi viene raccolto nelle vene che lo trasportano al cuore.
- La circolazione sanguigna termina nell'atrio.
Pertanto, sebbene i pesci non possano essere definiti animali a sangue caldo, i loro organi e tessuti ricevono sangue arterioso puro. Ciò aiuta i pesci a vivere nelle acque fredde dell'Artico e dell'Antartico e anche a non morire nei corpi d'acqua dolce in inverno.