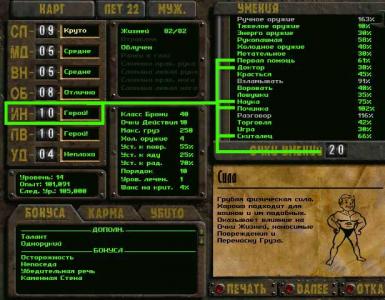
Sostanza intercellulare del tessuto nervoso. Tipi di tessuto nervoso. Centro e periferia
Il tessuto nervoso è il tessuto principale del sistema nervoso e le sue proprietà principali sono l'eccitabilità e la conduttività.
Il tessuto nervoso è costituito principalmente da cellule. Le sue cellule sono divise in 2 gruppi:
cellule nervose (neuroni): forniscono funzioni di conduzione ed eccitazione;
cellule neurogliali – forniscono funzioni ausiliarie (trofismo, protezione, ecc.)
2. Embriogenesi del tessuto nervoso.
La fonte embrionale del tessuto è il rudimento dell'ectoderma neurale, che forma il tubo neurale. Il tubo è costituito da 3 strati: interno (contiene cellule cambiali e dà origine alla glia ependimale); strato del mantello (mantello) (le cellule dello strato interno migrano qui e si differenziano in neuroblasti e ulteriormente in neuroni e spongioblasti, da cui si forma la maggior parte delle cellule neurogliali; velo marginale (contiene processi di cellule sottostanti).
3. Caratteristiche morfo-funzionali del neurone.
L'aspetto morfologico di un neurone corrisponde alle sue funzioni di eccitazione e conduzione degli impulsi nervosi, assicurata dal meccanismo di depolarizzazione delle membrane cellulari. Questo fenomeno si basa su una variazione della differenza di potenziale sulle superfici interna ed esterna delle membrane dovuta a correnti locali di Na+ nel citoplasma e di K+ fuori attraverso i canali ionici.
La cellula ha un corpo o pericario con un grande nucleo e processi situati al centro: dendriti (ce ne possono essere diversi e conducono l'eccitazione al corpo del neurone, ricevendolo attraverso numerosi contatti con altri neuroni. In queste aree si formano sporgenze speciali - spine dendritiche) e 1 assone (conduce l'eccitazione dal corpo al neurone o all'organo funzionante successivo). Ci sono tutti gli organelli di importanza generale (anche il centro della cellula). E ci sono strutture specifiche. Sostanza basofila i cui accumuli sono visibili nel pericario e nei dendriti, ma sono assenti nell'assone. Si tratta di densi accumuli di EPS granulare. Così come le neurofibrille, elementi citoscheletrici costituiti da neurofilamenti intermedi e microtubuli. Promuovono il trasporto di sostanze all'interno del neurone, che è particolarmente importante per i processi.
4. Sinapsi e loro classificazione.
I neuroni sono caratterizzati da un tipo speciale di contatti intercellulari: le sinapsi. La sinapsi chimica più caratteristica è tra la fine dell'assone e l'inizio del dendrite della cellula successiva. È costituito da: 1. parte presinaptica (assone) 2. fessura sinaptica 3. membrana postsinaptica (dendrite). L'estensione terminale dell'assone contiene vescicole sinaptiche con una sostanza speciale - un neurotrasmettitore, che viene prodotto nel corpo del neurone e viene rapidamente trasportato all'estensione assonale. L'eccitazione del primo neurone porta ad un rapido afflusso di calcio attraverso la fessura persinaptica nell'assone, che avvia l'esocitosi del neurotrasmettitore nella fessura sinaptica. La membrana postsinaptica contiene recettori che si legano al trasmettitore, provocandone la depolarizzazione e la formazione di un impulso nervoso, o iperpolarizzazione, provocando l'inibizione. Il trasmettitore eccitatorio è l'acetilcolina, il trasmettitore inibitorio è la glicina. Tieni presente che le sinapsi chimiche sono capaci solo di trasmissione unilaterale degli impulsi.
A seconda della posizione, le sinapsi possono essere axo-dendritiche, asso-somatiche e asso-assonali (inibitorie).
5. Classificazioni dei neuroni.
I neuroni sono classificati morfologicamente: in base al numero di processi.
Biochimicamente: secondo il mediatore rilasciato (ad esempio colinergico)
Funzionale: sensoriale, motoria, associativa.
Questa classificazione dipende da quale terminazione ha l'assone o il dendrite di un dato neurone, che è chiamata terminazione nervosa.
Nei neuroni sensoriali, i dendriti terminano con terminazioni nervose recettoriali specializzate nella percezione di stimoli esterni (esterocettori) o interni (interorecettori).
6. Terminazioni nervose sensibili.
Le terminazioni nervose sensibili si dividono in: libere e non libere. Quelli liberi sono semplicemente rami dendritici nell'epitelio o nel tessuto connettivo. Percepiscono segnali termici, meccanici e dolorosi.
Le terminazioni non libere sono non incapsulate o incapsulate. I primi sono rami dendritici circondati da speciali cellule neurogliali. Trovato nel derma e nelle mucose. Le terminazioni incapsulate non libere sono ricoperte anche esternamente da una capsula di tessuto connettivo. Questi includono una serie di meccanocettori che percepiscono la pressione e la vibrazione (corpuscoli lamellari di Vater-Pacini, corpuscoli tattili di Meissner, corpuscoli di Ruffini, ecc.), nonché fusi neuromuscolari: si tratta di recettori che si trovano all'interno dei muscoli scheletrici e valutano la grado di allungamento delle fibre muscolari I fusi contengono due tipi di fibre intrafusali: fibre a sacco nucleare e fibre a catena nucleare. Le terminazioni sensibili dei dendriti formano terminazioni a spirale ad anello e a forma di acino d'uva su queste fibre e rispondono ai cambiamenti nel loro spessore. Queste fibre hanno anche terminazioni motorie degli assoni, che le fanno contrarre al momento della contrazione dell'intero muscolo.
7. Terminazioni nervose efferenti.
Gli assoni dei motoneuroni formano terminazioni nervose effettrici di due tipi: secretorie (sulle cellule della ghiandola) e motorie (nei muscoli striati e lisci). Nel muscolo scheletrico è la giunzione neuromuscolare o placca motoria. La struttura è simile alla sinapsi che conoscete, ma la membrana postsinaptica è rappresentata da un tratto del plasmalemma della fibra muscolare. Un assone, ramificato all'estremità, forma contemporaneamente placche motorie su un intero gruppo di fibre muscolari. Nel tessuto cardiaco e muscolare liscio, i rami assonali formano estensioni - varicosità, in cui si trovano vescicole con il neurotrasmettitore. Di norma, solo alcune cellule sono innervate qui e da esse l'eccitazione viene trasmessa a quelle vicine con l'aiuto dei nessi.
Le terminazioni nervose secretorie terminano nelle vene varicose vicino alle cellule secretorie e stimolano la sintesi delle secrezioni o il processo di esocitosi.
8. Neuroglia.
Le neuroglia sono un gruppo di cellule di supporto che supportano l'attività dei neuroni. Nel tessuto cerebrale il loro numero è 5-10 volte maggiore di quello dei neuroni.
Ci sono microglia e macroglia. Le microglia sono piccole cellule stellate formate da monociti e sono macrofagi specializzati del sistema nervoso centrale. Svolgono una funzione protettiva, inclusa la funzione di presentazione dell'antigene. È stato chiarito il ruolo principale di queste cellule nei danni al sistema nervoso nell'AIDS. Diffondono il virus e avviano anche un aumento dell’apoptosi dei neuroni.
9. Caratteristiche e classificazione della macroglia.
La macroglia comprende diverse cellule appartenenti a tre varietà: astroglia, oligodendroglia e glia ependimale. Cellule gliali ependimali (ependimociti) Ependimociti.
Formano il rivestimento delle cavità dei ventricoli del cervello e del canale centrale del midollo spinale. Formano uno strato collegato da contatti intercellulari e giacciono sulla membrana basale, quindi sono anche classificati come epiteli. Separano i neuroni e il liquido cerebrospinale, formando una barriera neuro-liquor (altamente permeabile). E nell'area dei plessi coroidei fanno parte della barriera sangue-liquido cerebrospinale (tra il sangue e il liquido cerebrospinale). Questa barriera comprende: l'endotelio vascolare, la membrana che circonda i vasi, la membrana basale degli ependimociti e lo strato cellulare ependimale.
Gli oligodendroglia sono una varietà di piccole cellule con pochi e brevi processi che circondano i neuroni. Nei gangli racchiudono i corpi cellulari dei neuroni, fornendo una funzione di barriera. Un altro gruppo forma guaine lungo i processi dei neuroni, formando insieme a loro le fibre nervose. Nella periferia n.s. sono chiamati lemmociti o cellule di Schwann, nel sistema nervoso centrale - oligodendrociti.
Le astroglia sono rappresentate dagli astrociti: cellule stellate simili ai neuroni. Gli astrociti protoplasmatici sono caratteristici della sostanza grigia del sistema nervoso centrale e hanno processi corti e spessi; gli astrociti fibrosi sono caratteristici della sostanza bianca e hanno processi lunghi. Le loro funzioni sono di supporto (riempire gli spazi tra i neuroni), metabolica e regolatrice (mantenere una composizione costante di ioni e mediatori), barriera (parte della barriera emato-encefalica, che isola in modo affidabile i neuroni dal sangue, prevenendo il conflitto immunitario). La BBB comprende l'endotelio dei capillari e la loro membrana basale, nonché una densa guaina di processi astrocitari che ricopre i vasi.
10. Fibre nervose non mielinizzate e mielinizzate. Educazione e caratteristiche strutturali.
Le fibre nervose sono processi di neuroni (sono chiamati cilindri assiali), che sono ricoperti da una guaina di cellule gliali. Esistono fibre nervose mielinizzate e non mielinizzate.
Non mielinizzato le fibre si formano quando il cilindro assiale è immerso nei recessi dei lemmociti, che si trovano in una catena lungo l'intero assone. I lemmociti si piegano così tanto che le loro membrane si toccano sopra il cilindro assiale. Questa duplicazione è chiamata mesassone. Se più assoni sono immersi contemporaneamente in una catena di lemmociti, tale fibra viene chiamata cavo.
mielina fibre nervose. Si formano con la partecipazione delle cellule di Schwann, che prima formano un mesassone sopra il cilindro assiale, e poi iniziano a torcersi ripetutamente. Il citoplasma, insieme al nucleo, viene spinto verso l'esterno, formando uno strato chiamato neurolemma. Al di sotto di esso si trova uno spesso strato di doppie membrane strettamente adiacenti chiamate mielina. In alcune aree, tra le spire rimangono piccoli strati: tacche mieliniche. Perché le cellule di Schwann. L'assone è lungo e lungo di esso sono presenti numerose cellule di Schwann. Ai confini di due cellule vicine, la guaina mielinica scompare. Queste aree sono chiamate nodi di Ranvier.
Nel sistema nervoso centrale, la guaina mielinica è formata in modo leggermente diverso.
Le fibre mielinizzate conducono gli impulsi nervosi dieci volte più velocemente delle fibre non mielinizzate.
Tessuto nervoso controlla tutti i processi del corpo.
Il tessuto nervoso è costituito da neuroni(cellule nervose) e neuroglia(sostanza intercellulare). Le cellule nervose hanno forme diverse. Una cellula nervosa è dotata di processi ad albero - dendriti, che trasmettono stimoli dai recettori al corpo cellulare, e un lungo processo - un assone, che termina sulla cellula effettrice. A volte l'assone non è coperto da una guaina mielinica.
Le cellule nervose sono capaci sotto l'influenza dell'irritazione entrano in uno stato eccitazione, generare impulsi e trasmettere loro. Queste proprietà determinano la funzione specifica del sistema nervoso. Le neuroglia sono organicamente associate alle cellule nervose e svolgono funzioni trofiche, secretorie, protettive e di supporto.
Cellule nervose: i neuroni, o neurociti, sono cellule di processo. Le dimensioni del corpo del neurone variano ampiamente (da 3-4 a 130 micron). Anche le cellule nervose hanno una forma molto diversa. I processi delle cellule nervose conducono gli impulsi nervosi da una parte all'altra del corpo umano, la lunghezza dei processi va da diversi micron a 1,0-1,5 m.
Struttura del neurone. 1 - corpo cellulare; 2 - nucleo; 3 - dendriti; 4 - neurite (assone); 5 - estremità ramificata del neurite; 6 - neurilemma; 7 - mielina; 8 - cilindro assiale; 9 - intercettazioni di Ranvier; 10 - muscolo
Esistono due tipi di processi delle cellule nervose. I processi del primo tipo conducono gli impulsi dal corpo della cellula nervosa ad altre cellule o tessuti degli organi funzionanti; sono chiamati neuriti o assoni. Una cellula nervosa ha sempre un solo assone, che termina in un apparato terminale su un altro neurone o in un muscolo o una ghiandola. I processi del secondo tipo sono chiamati dendriti e si ramificano in un albero. Il loro numero varia tra i diversi neuroni. Questi processi conducono gli impulsi nervosi al corpo della cellula nervosa. I dendriti dei neuroni sensoriali hanno speciali dispositivi percettivi all'estremità periferica: terminazioni nervose sensoriali o recettori.
Classificazione dei neuroni per funzione:
- percepire (sensibile, sensoriale, recettore). Servono a percepire i segnali provenienti dall'ambiente esterno ed interno e trasmetterli al sistema nervoso centrale;
- contatto (intermedio, interneuroni, interneuroni). Fornire elaborazione, archiviazione e trasmissione di informazioni ai motoneuroni. Sono la maggioranza nel sistema nervoso centrale;
- motore (efferente). Generano segnali di controllo e li trasmettono ai neuroni periferici e agli organi esecutivi.
Tipi di neuroni per numero di processi:
- unipolare: avere un processo;
- pseudounipolare: un processo si estende dal corpo, che poi si divide in 2 rami;
- bipolare: due processi, uno dendrite, l'altro un assone;
- multipolare: ha un assone e molti dendriti.

Neuroni(cellule nervose). A - neurone multipolare; B - neurone pseudounipolare; B - neurone bipolare; 1 - assone; 2 - dendrite
Vengono chiamati gli assoni ricoperti da una guaina fibre nervose. Ci sono:
- continuo- ricoperti da una membrana continua, fanno parte del sistema nervoso autonomo;
- polposo- ricoperti da una membrana complessa e discontinua, gli impulsi possono spostarsi da una fibra ad altri tessuti. Questo fenomeno è chiamato irradiazione.

Terminazioni nervose. A - terminazione motoria su una fibra muscolare: 1 - fibra nervosa; 2 - fibra muscolare; B - terminazioni sensibili nell'epitelio: 1 - terminazioni nervose; 2 - cellule epiteliali
Terminazioni nervose sensoriali ( recettori) sono formati dai rami terminali dei dendriti dei neuroni sensoriali.
- esterocettori percepire le irritazioni provenienti dall'ambiente esterno;
- interorecettori percepire irritazioni dagli organi interni;
- propriocettori ricevere irritazioni dall'orecchio interno e dalle capsule articolari.
In base al loro significato biologico, i recettori si dividono in: cibo, sessuale, difensiva.
In base alla natura della risposta, i recettori si dividono in: il motore- si trovano nei muscoli; secretivo- nelle ghiandole; vasomotore-nei vasi sanguigni.
Effettore- collegamento esecutivo dei processi nervosi. Esistono due tipi di effettori: motori e secretori. Le terminazioni nervose motorie (motrici) sono i rami terminali dei neuriti delle cellule motorie nel tessuto muscolare e sono chiamate terminazioni neuromuscolari. Le terminazioni secretorie nelle ghiandole formano terminazioni neuroghiandolari. I tipi di terminazioni nervose menzionati rappresentano una sinapsi del tessuto nervoso.
La comunicazione tra le cellule nervose viene effettuata utilizzando le sinapsi. Sono formati dai rami terminali del neurite di una cellula del corpo, dai dendriti o dagli assoni di un'altra. In una sinapsi, un impulso nervoso viaggia in una sola direzione (da un neurite al corpo o ai dendriti di un'altra cellula). Sono disposti in modo diverso nelle diverse parti del sistema nervoso.
Il tessuto nervoso umano nel corpo ha diversi luoghi di localizzazione primaria. Questi sono il cervello (colonna vertebrale e testa), i gangli autonomi e il sistema nervoso autonomo (divisione metasimpatica). Il cervello umano è costituito da un insieme di neuroni, il cui numero totale supera il miliardo. Il neurone stesso è costituito da un soma - il corpo, nonché da processi che ricevono informazioni da altri neuroni - dendriti e un assone, che è una struttura allungata che trasmette informazioni dal corpo ai dendriti di altre cellule nervose.
Diversi tipi di processi nei neuroni
Il tessuto nervoso comprende un totale di fino a un trilione di neuroni di varie configurazioni. Possono essere unipolari, multipolari o bipolari a seconda del numero di processi. Le varianti unipolari con un processo sono rare negli esseri umani. Hanno un solo processo: l'assone. Questa unità del sistema nervoso è comune negli animali invertebrati (quelli che non possono essere classificati come mammiferi, rettili, uccelli e pesci). Vale la pena considerare che, secondo la classificazione moderna, fino al 97% di tutte le specie animali finora descritte sono classificate come invertebrati, quindi i neuroni unipolari sono abbastanza ampiamente rappresentati nella fauna terrestre.
Il tessuto nervoso con neuroni pseudounipolari (hanno un processo ma biforcato all'estremità) si trova nei vertebrati superiori nei nervi cranici e spinali. Ma più spesso, i vertebrati hanno campioni di neuroni bipolari (c'è sia un assone che un dendrite) o multipolari (un assone e diversi dendriti).

Classificazione delle cellule nervose
Quale altra classificazione ha il tessuto nervoso? I neuroni al suo interno possono svolgere diverse funzioni, quindi ce ne sono diversi tipi, tra cui:
- Anche le cellule nervose afferenti sono sensibili e centripete. Queste cellule sono di piccole dimensioni (rispetto ad altre cellule dello stesso tipo), hanno un dendrite ramificato e sono associate alle funzioni dei recettori di tipo sensoriale. Si trovano all'esterno del sistema nervoso centrale, hanno un processo situato in contatto con qualsiasi organo e un altro processo diretto nel midollo spinale. Questi neuroni creano impulsi sotto l'influenza dell'ambiente esterno o di eventuali cambiamenti nel corpo umano stesso. Le peculiarità del tessuto nervoso formato dai neuroni sensoriali sono tali che, a seconda del sottotipo di neuroni (monosensoriali, polisensoriali o bisensoriali), le reazioni possono essere ottenute sia rigorosamente a uno stimolo (mono) che a diversi (bi-, poli-) . Ad esempio, le cellule nervose nella zona secondaria della corteccia cerebrale (area visiva) possono elaborare sia stimoli visivi che uditivi. Le informazioni fluiscono dal centro alla periferia e ritorno.
- I neuroni motori (efferenti, motori) trasmettono informazioni dal sistema nervoso centrale alla periferia. Hanno un lungo assone. Il tessuto nervoso forma qui una continuazione dell'assone sotto forma di nervi periferici, che si avvicinano agli organi, ai muscoli (lisci e scheletrici) e a tutte le ghiandole. La velocità di eccitazione che passa attraverso l'assone nei neuroni di questo tipo è molto elevata.
- I neuroni intercalari (associativi) sono responsabili della trasmissione delle informazioni da un neurone sensoriale a quello motorio. Gli scienziati suggeriscono che il tessuto nervoso umano sia costituito dal 97-99% di tali neuroni. La loro localizzazione primaria è nella materia grigia del sistema nervoso centrale e possono essere inibitori o eccitatori a seconda delle funzioni che svolgono. I primi hanno la capacità non solo di trasmettere l'impulso, ma anche di modificarlo, aumentandone l'efficienza.

Gruppi specifici di cellule
Oltre alle suddette classificazioni, i neuroni possono essere attivi in background (le reazioni avvengono senza alcuna influenza esterna), mentre altri danno un impulso solo quando viene applicata loro una certa forza. Un gruppo separato di cellule nervose è costituito da neuroni rivelatori, che possono rispondere selettivamente ad alcuni segnali sensoriali che hanno significato comportamentale; sono necessari per il riconoscimento di schemi. Ad esempio, ci sono cellule nella neocorteccia che sono particolarmente sensibili ai dati che descrivono qualcosa di simile al volto di una persona. Le proprietà del tessuto nervoso qui sono tali che il neurone emette un segnale in qualsiasi posizione, colore, dimensione dello "stimolo facciale". Il sistema visivo contiene neuroni responsabili della rilevazione di fenomeni fisici complessi come oggetti che si avvicinano e si allontanano, movimenti ciclici, ecc.
Il tessuto nervoso in alcuni casi forma complessi molto importanti per il funzionamento del cervello, quindi alcuni neuroni hanno nomi personali in onore degli scienziati che li hanno scoperti. Si tratta di cellule Betz, di dimensioni molto grandi, che forniscono la comunicazione tra l'analizzatore motorio attraverso l'estremità corticale con i nuclei motori nel tronco cerebrale e un certo numero di parti del midollo spinale. Queste sono cellule inibitorie di Renshaw, al contrario, di piccole dimensioni, che aiutano a stabilizzare i motoneuroni quando si mantiene un carico, ad esempio sulla mano e a mantenere la posizione del corpo umano nello spazio, ecc.

Ci sono circa cinque neuroglia per ciascun neurone
La struttura del tessuto nervoso comprende un altro elemento chiamato “neuroglia”. Queste cellule, dette anche gliali o gliociti, hanno dimensioni 3-4 volte più piccole dei neuroni stessi. Nel cervello umano, ci sono cinque volte più neuroglia che neuroni, il che potrebbe essere dovuto al fatto che le neuroglia supportano i neuroni svolgendo varie funzioni. Le proprietà del tessuto nervoso di questo tipo sono tali che negli adulti i gliciti sono rinnovabili, a differenza dei neuroni, che non vengono ripristinati. Le “responsabilità” funzionali della neuroglia includono la creazione di una barriera emato-encefalica con l'aiuto degli astrociti gliali, che impediscono a tutte le grandi molecole, ai processi patologici e a molti farmaci di entrare nel cervello. I gliociti-olegodendrociti sono di piccole dimensioni e formano una guaina mielinica simile al grasso attorno agli assoni dei neuroni, che ha una funzione protettiva. La neuroglia fornisce anche supporto, trofica, delimitazione e altre funzioni.
Altri elementi del sistema nervoso
Alcuni scienziati includono anche l'ependima nella struttura del tessuto nervoso: un sottile strato di cellule che riveste il canale centrale del midollo spinale e le pareti dei ventricoli del cervello. Per la maggior parte, l'ependima è monostrato, costituito da cellule cilindriche, nel terzo e quarto ventricolo del cervello ha diversi strati. Le cellule che compongono l'ependima, gli ependimociti, svolgono funzioni secretorie, delimitanti e di sostegno. I loro corpi sono di forma allungata e hanno "ciglia" alle estremità, a causa del movimento del quale si muove il liquido cerebrospinale. Nel terzo ventricolo del cervello ci sono speciali cellule ependimali (taniciti), che si ritiene trasmettano dati sulla composizione del liquido cerebrospinale a una sezione speciale della ghiandola pituitaria.

Le cellule “immortali” scompaiono con l’età
Gli organi del tessuto nervoso, per definizione diffusa, comprendono anche le cellule staminali. Questi includono formazioni immature che possono diventare cellule di diversi organi e tessuti (potenza) e subire un processo di autorinnovamento. Infatti, lo sviluppo di qualsiasi organismo multicellulare inizia con una cellula staminale (zigote), dalla quale, attraverso la divisione e la differenziazione, si ottengono tutti gli altri tipi di cellule (nell'uomo ce ne sono più di duecentoventi). Lo zigote è una cellula staminale totipotente che dà origine ad un organismo vivente completo attraverso la differenziazione tridimensionale in unità tissutali extraembrionali ed embrionali (11 giorni dopo la fecondazione nell'uomo). I discendenti delle cellule totipotenti sono cellule pluripotenti, che danno origine agli elementi dell'embrione: endoderma, mesoderma ed ectoderma. È da quest'ultimo che si sviluppano il tessuto nervoso, l'epitelio cutaneo, le sezioni del tubo intestinale e gli organi di senso, quindi le cellule staminali sono parte integrante e importante del sistema nervoso.
Ci sono pochissime cellule staminali nel corpo umano. Ad esempio, un embrione ha una cellula del genere su 10mila, e una persona anziana di circa 70 anni ne ha una su cinque-otto milioni. Le cellule staminali, oltre alla potenza sopra menzionata, hanno proprietà come “homing” - la capacità di una cellula, dopo l'iniezione, di arrivare nell'area danneggiata e correggere i guasti, eseguendo le funzioni perdute e preservando i telomeri della cellula. In altre cellule, parte del telomero viene persa durante la divisione, ma nelle cellule tumorali, germinali e staminali esiste la cosiddetta attività telosize, durante la quale le estremità dei cromosomi vengono costruite automaticamente, il che offre infinite possibilità di divisione cellulare, cioè l'immortalità. Le cellule staminali, in quanto organi unici del tessuto nervoso, hanno un potenziale così elevato a causa dell'eccesso di informazioni sull'acido ribonucleico per tutti i tremila geni che partecipano alle prime fasi dello sviluppo dell'embrione.

Le principali fonti di cellule staminali sono gli embrioni, il materiale fetale dopo l'aborto, il sangue del cordone ombelicale, il midollo osseo, pertanto, dall'ottobre 2011, la decisione della Corte Europea vieta le manipolazioni con cellule staminali embrionali, poiché l'embrione è riconosciuto come persona da il momento della fecondazione. In Russia è consentito il trattamento con cellule staminali proprie e di donatori per una serie di malattie.
Sistema nervoso autonomo e somatico
I tessuti del sistema nervoso permeano tutto il nostro corpo. Dal sistema nervoso centrale (cervello, midollo spinale) partono numerosi nervi periferici che collegano gli organi del corpo con il sistema nervoso centrale. La differenza tra il sistema periferico e quello centrale è che non è protetto dalle ossa ed è quindi più facilmente suscettibile a lesioni varie. Secondo le sue funzioni, il sistema nervoso è diviso in sistema nervoso autonomo (responsabile dello stato interno di una persona) e sistema nervoso somatico, che entra in contatto con stimoli ambientali, riceve segnali senza trasferirli a fibre simili ed è controllato consapevolmente .
Vegetativo, d'altra parte, fornisce, piuttosto, l'elaborazione automatica e involontaria dei segnali in arrivo. Ad esempio, il dipartimento simpatico del sistema autonomo, quando il pericolo si avvicina, aumenta la pressione sanguigna, il polso e i livelli di adrenalina di una persona. Il dipartimento parasimpatico si attiva quando una persona riposa: le sue pupille si restringono, il battito cardiaco rallenta, i vasi sanguigni si dilatano e viene stimolato il funzionamento dei sistemi riproduttivo e digestivo. Le funzioni dei tessuti nervosi della parte enterica del sistema nervoso autonomo comprendono la responsabilità di tutti i processi digestivi. L'organo più importante del sistema nervoso autonomo è l'ipotalamo, che è associato alle reazioni emotive. Vale la pena ricordare che gli impulsi nei nervi autonomi possono divergere nelle fibre vicine dello stesso tipo. Pertanto, le emozioni possono influenzare chiaramente lo stato di una varietà di organi.
I nervi controllano i muscoli e altro ancora
I tessuti nervosi e muscolari nel corpo umano interagiscono strettamente tra loro. Pertanto, i principali nervi spinali (del midollo spinale) della regione cervicale sono responsabili del movimento dei muscoli alla base del collo (primo nervo) e forniscono il controllo motorio e sensoriale (2° e 3° nervo). Il nervo pettorale, che continua dal quinto, terzo e secondo nervo spinale, controlla il diaframma, supportando la respirazione spontanea.
I nervi spinali (dal quinto all'ottavo) si combinano con il nervo sternale per creare il plesso brachiale, che consente la funzione delle braccia e della parte superiore della schiena. La struttura del tessuto nervoso qui sembra complessa, ma è altamente organizzata e varia leggermente da persona a persona.
In totale, gli esseri umani hanno 31 paia di uscite dei nervi spinali, otto delle quali si trovano nella regione cervicale, 12 nella regione toracica, cinque ciascuna nella regione lombare e sacrale e una nella regione coccigea. Inoltre, ci sono dodici nervi cranici che provengono dal tronco cerebrale (la parte del cervello che continua il midollo spinale). Sono responsabili dell'olfatto, della vista, del movimento del bulbo oculare, del movimento della lingua, delle espressioni facciali, ecc. Inoltre, il decimo nervo qui è responsabile delle informazioni dal torace e dall'addome e l'undicesimo per il lavoro del trapezio e muscoli sternocleidomastoidei, che si trovano parzialmente all'esterno della testa. Tra i grandi elementi del sistema nervoso, vale la pena menzionare il plesso sacrale dei nervi, i nervi lombari, intercostali, i nervi femorali e il tronco del nervo simpatico.

Il sistema nervoso nel mondo animale è rappresentato da un'ampia varietà di campioni
Il tessuto nervoso degli animali dipende dalla classe a cui appartiene l'essere vivente in questione, sebbene i neuroni siano ancora una volta la base di tutto. Nella tassonomia biologica, un animale è considerato una creatura che ha un nucleo nelle sue cellule (eucarioti), è capace di movimento e si nutre di composti organici già pronti (eterotrofia). Ciò significa che possiamo considerare sia il sistema nervoso di una balena che, ad esempio, un verme. Il cervello di alcuni di questi ultimi, a differenza di quello umano, non contiene più di trecento neuroni, e il resto del sistema è un complesso di nervi attorno all'esofago. In alcuni casi, le terminazioni nervose che portano agli occhi sono assenti, poiché i vermi che vivono sottoterra spesso non hanno gli occhi.
Domande da considerare
Le funzioni dei tessuti nervosi nel mondo animale si concentrano principalmente sulla garanzia che il loro proprietario sopravviva con successo nell'ambiente. Allo stesso tempo, la natura nasconde molti misteri. Ad esempio, perché una sanguisuga ha bisogno di un cervello con 32 nodi nervosi, ognuno dei quali è di per sé un mini-cervello? Perché questo organo occupa fino all'80% dell'intera cavità corporea del ragno più piccolo del mondo? Ci sono anche evidenti sproporzioni nelle dimensioni dell'animale stesso e di parti del suo sistema nervoso. I calamari giganti hanno un "organo pensante" principale a forma di "ciambella" con un buco al centro e del peso di circa 150 grammi (con un peso totale fino a 1,5 centesimi). E tutto questo può essere oggetto di riflessione per il cervello umano.
Capitolo 10. TESSUTO NERVOSO
Capitolo 10. TESSUTO NERVOSO
Il tessuto nervoso è un sistema di cellule nervose interconnesse, neuroglia e macrofagi gliali, che forniscono funzioni specifiche di percezione delle irritazioni, eccitazione, generazione e trasmissione degli impulsi. È la base per la struttura degli organi del sistema nervoso. In ciascuna parte del sistema nervoso, la composizione cellulare-differenziale del tessuto nervoso e le sue caratteristiche morfofunzionali sono uniche. Ciò garantisce la regolazione ottimale delle funzioni vitali di tutti i tessuti e gli organi, la loro integrazione nel corpo e il collegamento con l'ambiente.
Cellule nervose(neuroni, neurone)- i principali elementi istologici del tessuto nervoso, che percepiscono il segnale e lo trasmettono ad altre cellule nervose o cellule effettrici con l'aiuto di neurotrasmettitori. Neuroglia garantisce l'esistenza e il funzionamento delle cellule nervose, svolge funzioni di sostegno, trofiche, delimitanti, secretorie e protettive. Microglia- cellule, alcune delle quali appartengono al sistema dei fagociti mononucleari (vedi sotto).
10.1. SVILUPPO DEL TESSUTO NERVOSO
Il tessuto nervoso si sviluppa dalla parte dorsale dell'ectoderma. In un embrione umano di 18 giorni, l’ectoderma lungo la linea mediana dorsale si differenzia e si ispessisce, formando placca neurale, i cui bordi laterali si alzano, formandosi pieghe nervose, e tra le creste si forma un nervo scanalatura. L'estremità anteriore della placca neurale si espande, formando successivamente il cervello. I bordi laterali continuano a salire e crescere medialmente finché non si incontrano e si fondono lungo la linea mediana tubo neurale che è separato dall'ectoderma epidermico sovrastante. La cavità del tubo neurale persiste negli adulti come sistema ventricolare del cervello e canale centrale del midollo spinale. Alcune cellule della placca neurale non fanno parte del tubo neurale ed epi-
Riso. 10.1. Formazione del tubo neurale dell'embrione di pulcino (secondo A. G. Knorre): UN- stadio della placca neurale; B- chiusura del tubo neurale; V- separazione del tubo neurale e della placca gangliare dall'ectoderma. 1 - solco neurale; 2 - pieghe neurali; 3 - ectoderma cutaneo; 4 - accordo; 5 - mesoderma; 6 - piastra gangliare; 7 - tubo neurale; 8 - mesenchima; 9 - endoderma
ectoderma dermico e formano grappoli sui lati del tubo neurale, che si fondono in un cordone lasso situato tra il tubo neurale e l'ectoderma epidermico, - cresta neurale(placca gangliare) (Fig. 10.1). Dal tubo neurale si formano successivamente i neuroni e la neuroglia (macroglia) del sistema nervoso centrale. La cresta neurale dà origine ai neuroni dei gangli sensibili (sensoriali) e autonomici, alle cellule della pia madre e della membrana aracnoidea del cervello e ad alcuni tipi di glia: neurolemmociti (cellule di Schwann), cellule satellite gangliari
glia, cellule della midollare del surrene, melanociti della pelle, parte delle cellule del sistema endocrino disperso, cellule sensoriali dei corpi carotidei, ecc.
Nella formazione dei gangli dei nervi cranici V, VII, IX e X, oltre alla cresta neurale, anche i placodi neurali (neurogenici), che sono ispessimenti dell'ectoderma sui lati del tubo neurale in formazione nella regione cranica dell'embrione, prendono parte anche loro.
Il tubo neurale nelle prime fasi dell'embriogenesi è una fila multipla neuroepitelio, costituito da steli matrice cellule (ventricolari). Le cellule della matrice, come risultato della mitosi asimmetrica e sotto l'influenza di fattori microambientali, sono capaci di differenziazione divergente in vari differenziamenti cellulari - neuroblastico, glioblastico E ependimoblastico. Un aumento del numero di cellule porta alla formazione di quattro zone concentriche nel tubo neurale, limitate dalle membrane limitanti gliali superficiali e periventricolari: ventricolare (ependimale), subventricolare, intermedia (mantello) e marginale (marginale) (Fig. 10.2 , UN).
Ventricolare La zona (ependimale) è costituita da cellule staminali (matrice) in divisione di forma cilindrica. Il nucleo della cellula ventricolare migra verso quella parte della cellula che si affaccia sul canale centrale. Le cellule si dividono e, dopo la divisione, i nuclei delle cellule figlie si spostano nelle parti apicali delle cellule risultanti, dove avviene nuovamente la replicazione del DNA. Il ciclo mitotico e il ciclo di migrazione nucleare durano dalle 5 alle 24 ore.
Subventricolare la zona è costituita da cellule che hanno perso la capacità di muovere i nuclei, ma mantengono un'elevata attività proliferativa. La zona subventricolare viene determinata nella regione del midollo spinale entro diversi giorni, ma in quelle aree del cervello in cui l'istogenesi avviene in modo particolarmente intenso, si formano zone germinali (cambiali) subventricolari ed extraventricolari che esistono da molto tempo. Pertanto, la zona cambiale extraventricolare del cervelletto scompare nell'uomo entro 20 mesi dall'ontogenesi postnatale.
Intermedio(mantello, mantello) è costituita da cellule che si sono spostate dalle zone ventricolari e subventricolari: neuroblasti e glioblasti. I neuroblasti perdono la capacità di dividersi e successivamente di differenziarsi in neuroni. I glioblasti continuano a dividersi e danno origine ad astrociti e oligodendrociti. Le forme mature di quest'ultimo non perdono completamente la capacità di dividersi. Poiché il numero di neuroni nel cervello è di circa 1 trilione, sembra che in media si formino 2.500.000 neuroni durante l'intero periodo prenatale di 1 minuto. Le cellule della zona intermedia formano la materia grigia del midollo spinale e parte della materia grigia del cervello.
Marginale La zona (velo marginale) è formata da assoni di neuroblasti e macroglia che crescono al suo interno e dà origine alla sostanza bianca. In alcune aree del cervello, le cellule della zona intermedia migrano ulteriormente, formando placche corticali - gruppi di cellule da cui si formano la corteccia cerebrale e il cervelletto.

Riso. 10.2. Sviluppo del cervello e differenziazione neuronale:
UN- midollo spinale a diversi stadi di sviluppo (secondo Hardesty); I - placca neurale, II, III - tubo neurale nelle fasi successive di sviluppo: 1 - cellula che si divide mitoticamente della placca neurale; 2 - cellula che si divide mitoticamente nella zona ventricolare (strato ependimale); 3 - zona intermedia (nucleare, strato di mantello); 4 - zona marginale (strato esterno, velo marginale); 5 - membrana limitante interna; 6 - membrana limitante esterna; 7 - mesenchima; B- stadi di differenziazione del neurone efferente del ganglio intracardiaco umano (secondo V.N. Shvalev, A.A. Sosunov, G. Guski): I - neuroblasto; II - neuroblasto con processi in via di sviluppo; III - neurone giovane con formazione di vescicole sinaptiche e sinapsi; IV - neurone differenziante con organelli nel pericario e un assone in crescita; V - neurone maturo con un grande pericario, numerose sinapsi e un assone che ha formato una terminazione neuromuscolare sui cardiomiociti; A 1 - fibre pregangliari; A 2 - fibre postgangliari; Eff - terminazione neuromuscolare efferente; AS - sinapsi asso-somatiche; AD - sinapsi assodendritiche; G - gliociti
Nella differenziazione neuroblastica, oltre alle cellule della matrice, sono presenti ney-roblasts, giovane E neuroni maturi. Rispetto alle cellule della matrice, la struttura ultramicroscopica del nucleo e del citoplasma cambia nel neuroblasto. Aree di diversa densità elettronica appaiono nel nucleo sotto forma di piccoli granelli e fili. Nel citoplasma, i tubuli e le cisterne del reticolo endoplasmatico granulare si rivelano in gran numero, il numero di ribosomi e polisomi liberi diminuisce, il complesso del Golgi raggiunge uno sviluppo significativo, vengono rilevate fibrille sottili - fasci di neurofilamenti e microtubuli. Il numero di neurofilamenti contenenti proteine - la tripletta di neurofilamenti - aumenta nel processo di specializzazione. Il corpo del neuroblasto acquisisce gradualmente una forma a forma di pera e dalla sua estremità appuntita inizia a svilupparsi un processo: assone(neurite). Successivamente altri processi si differenziano: dendriti.
Un neurone giovane è caratterizzato da un aumento del volume cellulare, dalla crescita dei processi, dalla formazione di una sostanza cromatofila e dalla comparsa delle prime sinapsi.
Nel processo di differenziazione dei neuroni dai neuroblasti si distinguono i periodi pre-mediatore e mediatore (vedi Fig. 10.2, B). Il periodo pre-mediatore è caratterizzato dal graduale sviluppo degli organelli di sintesi nel corpo dei ribosomi liberi dai neuroblasti e quindi dal reticolo endoplasmatico. Nel periodo del mediatore, le prime vescicole contenenti un mediatore compaiono nei neuroni giovani, e nei neuroni maturi c'è uno sviluppo significativo degli organelli di sintesi e secrezione (reticolo endoplasmatico granulare, complesso di Golgi), accumulo di mediatori e il loro ingresso nell'assone, e la formazione delle sinapsi. In generale, lo sviluppo di un neurone maturo è il processo più lungo. La cellula assume la sua forma finale, l'organizzazione istochimica, si integra nell'arco riflesso e nella rete neurale, si stabiliscono relazioni neuronali-gliali, ecc.
Nonostante la formazione del sistema nervoso sia completata nei primi anni dello sviluppo postnatale, la plasticità del sistema nervoso centrale rimane fino alla vecchiaia. Questa plasticità può essere espressa nella comparsa di nuovi terminali e di nuove connessioni sinaptiche. I neuroni del sistema nervoso centrale dei mammiferi sono in grado di formare nuovi rami (gemmazione assonale) e nuove sinapsi (sostituzione sinaptica). La plasticità si manifesta maggiormente nei primi anni dopo la nascita, ma persiste parzialmente negli adulti, ad esempio quando i livelli ormonali cambiano, quando si imparano nuove abilità o quando si verificano infortuni. Sebbene i neuroni siano permanenti, le loro connessioni sinaptiche possono essere modificate nel corso della vita, il che può esprimersi, in particolare, in un aumento o una diminuzione del loro numero. La plasticità nel danno cerebrale minore si manifesta con il ripristino parziale delle funzioni.
Nella popolazione dei neuroni, a partire dalle prime fasi di sviluppo del sistema nervoso e durante l'ontogenesi, si verifica una morte cellulare massiccia, che raggiunge il 25-75% dell'intera popolazione. Questa morte cellulare fisiologica programmata (apoptosi) avviene sia nel sistema nervoso centrale che periferico; in questo caso il cervello perde circa lo 0,1% dei neuroni.
10.2. NEURONI
Neurone (neurone), o cellula nervosa, è una cellula specializzata del sistema nervoso responsabile della percezione, dell'elaborazione degli stimoli, della conduzione degli impulsi e dell'influenza su altri neuroni, cellule muscolari o secretrici. I neuroni secernono neurotrasmettitori e altre sostanze che trasmettono informazioni. Un neurone è un'unità morfologicamente e funzionalmente indipendente, ma con l'aiuto dei suoi processi stabilisce un contatto sinaptico con altri neuroni, formando archi riflessi- anelli della catena da cui è costruito il sistema nervoso. A seconda della funzione nell'arco riflesso, ci sono recettore(sensibile, afferente), associativo E efferente Neuroni (effettori). I neuroni afferenti percepiscono l'impulso, i neuroni efferenti lo trasmettono ai tessuti degli organi funzionanti, spingendoli all'azione, ei neuroni associativi comunicano tra neuroni. I neuroni sono disponibili in un’ampia varietà di forme e dimensioni. Il diametro dei corpi cellulari dei granuli della corteccia cerebellare è di 4-6 µm e i neuroni piramidali giganti della zona motoria della corteccia cerebrale sono di 130-150 µm. Tipicamente i neuroni sono costituiti da corpi, O perikarya (corpo dei neuroni), e spara: assone e vari numeri di ramificazioni dendriti. Dal numero di processi che distinguono unipolare neuroni che hanno solo un assone (di solito non si trovano negli animali superiori e negli esseri umani), bipolare, avente un assone e un dendrite, e multipolare, avente un assone e molti dendriti (Fig. 10.3, 10.4). A volte tra le persone bipolari
Riso. 10.3. Neurone (secondo I.F. Ivanov): 1 - corpo del neurone; 2 - cilindro assiale; 3 - guaina mielinica in sezione; 4 - nuclei di neurolemmociti; 5 - strato di mielina; 6 - tacca mielinica; 7 - intercettazione nodale della fibra nervosa; 8 - fibra nervosa priva di mielina; 9 - terminazione neuromuscolare (motoria); 10 - fibre nervose mielinizzate trattate con acido osmico

Riso. 10.4. Tipi di cellule nervose (secondo T. N. Radostina, L. S. Rumyantseva):
UN- neurone unipolare; B- neurone pseudounipolare; V- neurone bipolare;
G- neurone multipolare
Ronov si incontra pseudounipolare, dal corpo di cui si estende una crescita comune - un processo, che poi si divide in un dendrite e un assone (Fig. 10.4, b). Nei gangli spinali sono presenti neuroni pseudounipolari, negli organi sensoriali sono presenti neuroni bipolari. La maggior parte dei neuroni sono multipolari. La loro forma è estremamente varia. L'assone e i suoi collaterali terminano ramificandosi in diversi rami chiamati telodendri, questi ultimi terminano con ispessimenti terminali.
La regione tridimensionale in cui vengono chiamati i dendriti di un ramo neuronale campo dendritico.
Citoplasma di un neurone. La stragrande maggioranza dei neuroni umani contiene un nucleo, il più delle volte situato al centro, meno spesso - eccentricamente. I neuroni binucleari e, soprattutto, multinucleati sono estremamente rari. Un'eccezione sono i neuroni di alcuni gangli del sistema nervoso autonomo; per esempio, nella prostata e nella cervice si trovano talvolta neuroni contenenti fino a 15 nuclei. La forma dei nuclei dei neuroni è rotonda. In conformità con l'elevata attività metabolica dei neuroni, la cromatina nei loro nuclei ha un basso grado di condensazione. Il nucleo ha uno, e talvolta due o tre nucleoli grandi. Un aumento dell'attività funzionale dei neuroni è solitamente accompagnato da un aumento del volume (e del numero) dei nucleoli.
Sostanza cromatofila(corpi tigroidi o di Nissl). Quando il tessuto nervoso viene colorato con coloranti all'anilina (tionina, blu di toluidina, viola cresilico, ecc.), nel citoplasma dei neuroni viene rilevata una sostanza cromatofila sotto forma di grumi e granuli basofili di varie dimensioni e forme. (sostantia cromatofila)(vedi Fig. 10.5, a). Basofilo

Riso. 10.5. Sostanza cromatofila (grumi di Nissl) e apparato neurofibrillare nei neuroni (micropreparati). La struttura di un neurone secretorio (secondo I. G. Akmaev): UN- sostanza cromatofila (colorata con blu di toluidina secondo il metodo Nissl); B- neurofibrille; V- neurone unipolare (b, V- impregnazione con nitrato d'argento): 1 - sostanza cromatofila; 2 - assone; 3 - dendriti; 4 - processo neuronale; G- neurone secretorio: 1 - nucleo; 2 - tubuli del reticolo endoplasmatico; 3 - grappoli di tubuli; 4 - Complesso del Golgi; 5 - granuli neurosecretori; 6 - mitocondri; 7 - lisosomi; 8 - sinapsi; 9 - emocapillare; 10 - epitelio ependimale dei ventricoli cerebrali; 11 - lobo anteriore della ghiandola pituitaria

Riso. 10.6. Organizzazione ultrastrutturale di una cellula nervosa nella corteccia cerebrale (schema secondo I. G. Pavlova):
1 - plasmalemma; 2 - nucleo; 3 - reticolo endoplasmatico granulare (sostanza cromatofila); 4 - Complesso del Golgi; 5 - lisosomi; 6 - mitocondri; 7 - neurofilamenti; 8 - microtubuli; 9 - dendrite; 10 - sinapsi assodendritiche;
11 - sinapsi assosomatiche
i grumi sono localizzati nel perikarya e nei dendriti dei neuroni, ma non si trovano mai negli assoni e nelle loro basi a forma di cono - collinette assonali. La basofilia dei grumi è spiegata dall'alto contenuto di ribonucleoproteine. La microscopia elettronica ha mostrato che ogni ciuffo di sostanza cromatofila è costituito da cisterne del reticolo endoplasmatico granulare, ribosomi liberi e polisomi (Fig. 10.6).
Il reticolo endoplasmatico granulare sintetizza proteine neurosecretrici, proteine integrali della membrana plasmatica e proteine lisosomiali. I ribosomi e i polisomi liberi sintetizzano proteine citosoliche (ialoplasma) e proteine non integrali del plasmalemma dei neuroni. I neuroni richiedono enormi quantità di proteine per svolgere le loro funzioni. Caratteristica degli assoni
flusso costante di citoplasma dal pericario ai terminali ad una velocità di 1-3 mm al giorno.
Il complesso del Golgi nei neuroni è ben sviluppato. Al microscopio ottico si rivela sotto forma di anelli, fili ritorti e grani di varie forme. La sua ultrastruttura è normale. Le vescicole del complesso di Golgi trasportano le proteine sintetizzate nel reticolo endoplasmatico granulare al plasmalemma (proteine integrali), o ai terminali (neuropeptidi, neurosecrezione), o ai lisosomi (idrolasi lisosomiali e membrane lisosomiali).
I mitocondri forniscono energia per processi come il trasporto degli ioni e la sintesi proteica. I neuroni necessitano di un apporto costante di glucosio e ossigeno dal sangue e l’interruzione dell’afflusso di sangue al cervello provoca la perdita di coscienza.
I lisosomi sono coinvolti nella degradazione enzimatica dei componenti cellulari, dei recettori e delle membrane.
Degli elementi citoscheletrici nel citoplasma dei neuroni ci sono filamenti intermedi (neurofilamenti) con un diametro di 12 nm, microtubuli (neurotubuli) con un diametro di 24-27 nm e microfilamenti di actina. Fasci di neurofilamenti su preparati impregnati di nitrato d'argento sono visibili al microscopio ottico sotto forma di fili - neurofibrille, che sono essenzialmente un artefatto (vedi Fig. 10.5, avanti Cristo). I microtubuli e le proteine associate forniscono il trasporto citoplasmatico delle sostanze, soprattutto nell'assone. I filamenti intermedi svolgono una funzione meccanica, mantenendo la forma del corpo e dei processi del neurone. I filamenti di actina, insieme ad altre proteine, sono coinvolti nel cambiamento della forma del corpo e dei processi del neurone (ad esempio nel cono di crescita).
Dendriti sono processi ramificati che iniziano con i recettori. Nella loro parte prossimale contengono gli stessi organelli del corpo cellulare: ciuffi di sostanza cromatofila, mitocondri, un gran numero di neurotubuli (microtubuli) e neurofilamenti. Il plasmalemma dei dendriti ha dei recettori, a seguito dei quali conducono l'eccitazione al pericario. A causa dei dendriti, la superficie recettoriale di un neurone aumenta di 1000 volte o più. Pertanto, i dendriti dei neuroni piriformi (cellule di Purkinje) della corteccia cerebellare aumentano la superficie del recettore da 250 a 27.000 μm 2 e sulla superficie di queste cellule si trovano fino a 200.000 terminazioni sinaptiche. I dendriti di molti neuroni hanno piccole proiezioni - spine. Si tratta di strutture dinamiche che possono cambiare forma e dimensione, il che influenza la trasmissione sinaptica degli impulsi nervosi al corpo del neurone.
Assone- un processo attraverso il quale l'impulso viene trasmesso dal corpo cellulare. La sua lunghezza varia da pochi micrometri a un metro. L'assone contiene mitocondri, neurotubuli e neurofilamenti, reticolo endoplasmatico e corpi multivescicolari con un diametro di circa 0,5 μm. Viene chiamata la posizione in cui l'assone ha origine dal corpo del neurone collinetta dell'assone. Questo è il sito di generazione del potenziale d'azione. Un gran numero di canali ionici si trovano qui nel plasmalemma. I canali ionici possono essere aperti, chiusi o

Riso. 10.7. Zone funzionali del neurone multipolare (secondo G. R. Nobak, N. L. Strominger, R. Demarest):
I - segmento del recettore; II - segmento trasmittente; III - segmento effettore. 1 - corpo del neurone con nucleo; 2 - dendriti; 3 - assone con guaina mielinica; 4 - fibra muscolare con terminali assonici; 5 - cambiamenti nel potenziale di membrana
inattivato. In un neurone a riposo, il potenziale di membrana a riposo è 60-70 mV. Il potenziale di riposo viene creato dalla rimozione di Na+ dalla cellula. La maggior parte dei canali Na+ e K+ sono chiusi. La transizione dei canali dallo stato chiuso a quello aperto è regolata dal potenziale di membrana (Fig. 10.7).
Come risultato dell'arrivo di un impulso eccitante al plasmalemma della cellula, si verifica una depolarizzazione parziale. Quando raggiunge un livello critico (soglia), i canali del sodio si aprono, consentendo agli ioni Na+ di entrare nella cellula. La depolarizzazione aumenta e, allo stesso tempo, si aprono ancora più canali del sodio. Potrebbe anche esserci peripolarizzazione, un potenziale di membrana inverso, quando la superficie esterna della membrana plasmatica è caricata negativamente e quella rivolta verso il citoplasma è caricata positivamente. I canali del sodio vengono inattivati entro 1-2 ms. Anche i canali del potassio si aprono, ma più lentamente e per un periodo di tempo più lungo, il che consente al K+ di lasciare la cellula e ripristinare il potenziale al livello precedente, altrimenti può verificarsi un'iperpolarizzazione. Dopo 1-2 ms (periodo refrattario), i canali ritornano al loro stato normale e la membrana può nuovamente rispondere agli stimoli. Quindi, la propagazione del potenziale d'azione è causata dall'ingresso di ioni Na + nel neurone, che possono depolarizzare l'area vicina del plasmalemma, che a sua volta crea un potenziale d'azione in un nuovo posto. Le caratteristiche della trasmissione degli impulsi nervosi nelle fibre nervose mielinizzate verranno delineate dopo aver descritto la loro struttura.
Trasporto assonale(trasporto assoplasmatico) è il movimento delle sostanze dal corpo ai processi e dai processi al corpo del neurone. È diretto dai tubuli neuronali; le proteine - chinesina e dineina - sono coinvolte nel trasporto. Il trasporto di sostanze dal corpo cellulare ai processi è chiamato anterogrado, al corpo retrogrado. Il trasporto assonale è rappresentato da due componenti principali: una componente veloce (400-2000 mm al giorno) ed una componente lenta (1-2 mm al giorno). Entrambi i sistemi di trasporto sono presenti sia negli assoni che nei dendriti.
Il sistema veloce anterogrado conduce strutture membranose, inclusi componenti della membrana, mitocondri, vescicole contenenti peptidi, precursori dei neurotrasmettitori e altre proteine. Il sistema veloce retrogrado conduce i materiali usati per la degradazione nei lisosomi, la distribuzione e il riciclaggio e possibilmente i fattori di crescita nervosa. I neurotubuli sono organelli responsabili del trasporto rapido, detti anche neurotubulosi-dipendenti. Quando i tubuli neuronali vengono distrutti, il trasporto rapido si interrompe. ATP e Ca 2+ forniscono questi movimenti. Su un tubo neuronale, le vescicole possono superare altre vescicole che si muovono nella stessa direzione. Allo stesso tempo, due vescicole possono muoversi in direzioni opposte lungo un tubo neuronale. Il trasporto lento è un sistema anterogrado che conduce proteine e altre sostanze per rinnovare e mantenere l'assoplasma (citosol) dei neuroni maturi e fornire assoplasma per la crescita di assoni e dendriti durante lo sviluppo e la rigenerazione.
Il trasporto assonale è un'espressione dell'unità dei neuroni. Grazie ad esso viene mantenuta una connessione costante tra il corpo cellulare (centro trofico) e i processi. Con il suo aiuto il corpo cellulare viene informato sui bisogni metabolici e sulle condizioni delle parti distali. Attraverso l'assorbimento di sostanze extracellulari, come il fattore di crescita nervoso, seguito dal trasporto retrogrado, il corpo cellulare può "valutare" il suo ambiente. Tuttavia, il trasporto retrogrado ha una proprietà negativa. Con esso, i virus neurotropi, come il virus della rabbia, vengono consegnati al sistema nervoso centrale. Tubuli neuronali difettosi possono essere la causa di alcuni disturbi neurologici negli esseri umani.
Neuroni secretori
La capacità di sintetizzare e secernere sostanze biologicamente attive, in particolare mediatori (acetilcolina, norepinefrina, serotonina, ecc.), è caratteristica di tutti i neuroni. Esistono però neuroni specializzati principalmente per svolgere questa funzione: neuroni secretori (neuronum secretorium), per esempio, le cellule dei nuclei neurosecretori della regione ipotalamica del cervello (vedi Fig. 10.5, d). I neuroni secretori hanno una serie di caratteristiche morfologiche specifiche. Questi sono neuroni grandi. La sostanza cromatofila si trova prevalentemente alla periferia del corpo cellulare. Nel citoplasma dei neuroni e negli assoni sono presenti granuli di secrezione di varie dimensioni - neurosecretoria (sostantia neurosecretoria), contenenti proteine e in alcuni casi lipidi e poli-
saccaridi. I granuli di neurosecrezione vengono rilasciati nel sangue o nel liquido cerebrospinale. Molti neuroni secretori hanno nuclei di forma irregolare, che indica la loro elevata attività funzionale. I neurosegreti giocano un ruolo neuroregolatori, partecipando all'interazione dei sistemi di integrazione nervosa e umorale.
10.3. NEUROGLIA
I neuroni sono cellule altamente specializzate che esistono e funzionano in un ambiente strettamente definito. Fornisce loro un tale ambiente neuroglia (neuroglia). Le neuroglia svolgono le seguenti funzioni: sostegno, trofica, delimitazione, mantenimento di un ambiente costante attorno ai neuroni, protettiva, secretiva. Ci sono glia centrale E sistema nervoso periferico(Fig. 10.8-10.10).

Riso. 10.8. Gliociti di vario tipo (secondo T. N. Radostina e L. S. Rumyantseva):
1 - ependimociti; 2 - astrociti protoplasmatici; 3 - astrozia fibrosa-
Voi; 4 - oligodendrociti; 5 - microglia
Glia del sistema nervoso centrale. Le cellule gliali del sistema nervoso centrale sono divise in macroglia(gliociti) e microglia. La macroglia si sviluppa dai glioblasti del tubo neurale. La macroglia comprende ependimociti, astrociti e oligodendrociti.
10.3.1. Macroglia
Ependimociti(ependimociti) allineano i ventricoli del cervello e il canale centrale del midollo spinale (Fig. 10.11). Queste sono cellule cilindriche. Formano uno strato come l'epitelio. Ci sono giunzioni e bande adesive tra le cellule adiacenti, ma non ci sono giunzioni strette, in modo che il liquido cerebrospinale possa penetrare tra di loro nel tessuto nervoso. La maggior parte degli ependimociti ha ciglia mobili che provocano il flusso del liquido cerebrospinale. La superficie basale della maggior parte degli ependimociti è liscia, ma alcune cellule hanno un lungo processo che si estende in profondità nel tessuto nervoso e sono quasi prive di ciglia. Tali cellule sono chiamate tanyciti. Si trovano in grandi quantità nel pavimento del terzo ventricolo. Si ritiene che queste cellule trasmettano informazioni sulla composizione del liquido cerebrospinale alla rete capillare primaria del sistema portale ipofisario. L'epitelio ependimale dei plessi corioidei dei ventricoli produce liquido cerebrospinale. Il citoplasma degli ependimociti contiene numerosi mitocondri, il complesso del Golgi situato sopra il nucleo e un reticolo endoplasmatico granulare poco sviluppato.

Riso. 10.9. Partecipazione dei gliciti alla formazione delle fibre mieliniche nella parte centrale (a) e periferica (B) sistema nervoso (secondo K. L. Junqueira, H. Carneiro, P. O. Keley):
1 - dendriti; 2 - sinapsi; 3 - perika-rion; 4 - collinetta dell'assone; 5 - assone; 6 - mielina; 7 - oligodendrociti; 8 - intercettazione nodale; 9 - neurolemmociti (cellule di Schwann); 10 - giunzione neuromuscolare
Astrociti(astrociti, dal greco astron- stella, Kyoto- cellula) - cellule a forma di processo, povere di organelli. Eseguono principalmente il supporto, la delimitazione

Riso. 10.10. Rapporti tra neuroni, astroglia, oligodendroglia e terminali nervosi (secondo G. R. Nobak, N. L. Strominger, R. D. Demarest):
1 - corpo del neurone; 2 - dendriti; 3 - assone; 4 - astrocito; 5 - oligodendrociti; 6 - sinapsi assoassonale; 7 - sinapsi assodendritica; 8 - sinapsi assosomatica; 9 - capillare; 10 - piede perivascolare dell'astrocito

Riso. 10.11. Ependimociti del ventricolo cerebrale (secondo G. R. Nobak, G. L. Strominger, R. D. Demarest):
1 - cavità ventricolare; 2 - ependimociti; 3 - capillari del plesso coroideo; 4 - cervello; 5 - membrana morbida del cervello; 6 - membrana aracnoidea; 7 - spazio subaracnoideo; 8 - neuroni

Riso. 10.12. Oligodendrociti e formazione di strati di mielina nelle fibre nervose del sistema nervoso centrale (secondo Bunge et al.):
1 - oligodendrociti; 2 - fibre nervose; 3 - citoplasma degli oligodendrociti; 4 - assone; 5 - spazio intercellulare
funzioni corporee e metaboliche (vedi Fig. 10.10). Ci sono astrociti protoplasmatici (astrociti protoplasmatici), localizzato nella materia grigia del sistema nervoso centrale e negli astrociti fibrosi (fibrosi astrocitica), presente nella sostanza bianca.
Astrociti protoplasmatici caratterizzato da processi corti e molto ramificati e da un nucleo sferico leggero. Astrociti fibrosi hanno 20-40 processi lunghi, debolmente ramificati, in cui sono presenti molte fibrille costituite da filamenti intermedi di diametro
10 miglia nautiche Nei filamenti viene rilevata la proteina acida fibrillare gliale. I processi degli astrociti si estendono alle membrane basali dei capillari, ai corpi e ai dendriti dei neuroni, circondano le sinapsi e le separano l'una dall'altra (vedi Fig. 10.8, 10.12), nonché alla pia madre del cervello, formando membrana limitante piogliale, confinante con lo spazio subaracnoideo. Avvicinandosi ai capillari, i loro processi formano "gambe" espanse che circondano completamente la nave. Gli astrociti accumulano e trasferiscono sostanze dai capillari ai neuroni, assorbendo il potassio extracellulare in eccesso e altre sostanze, come i neurotrasmettitori, dallo spazio extracellulare dopo un'intensa attività neuronale.
Oligodendrociti(oligodendrociti hanno nuclei più piccoli e più intensamente colorati rispetto agli astrociti. I loro processi sono pochi. Gli oligodendrociti sono presenti sia nella sostanza grigia che in quella bianca. Nella materia grigia sono localizzati vicino al pericarya. Nella sostanza bianca, i loro processi partecipano alla formazione dello strato mielinico nelle fibre nervose mielinizzate e, a differenza dei neurolemmociti del sistema nervoso periferico, un oligodendrocita può partecipare alla mielinizzazione di diversi assoni (vedi Fig. 10.8, Fig. 10.12).
Un processo forma lo strato mielinico di un segmento internodale. Il citoplasma degli oligodendrociti è elettrodenso, contiene molti mitocondri, un complesso di Golgi ben sviluppato, cisterne del reticolo endoplasmatico granulare e numerosi microtubuli.
10.3.2. Microglia
La popolazione microgliale è di origine eterogenea. Circa la metà delle cellule microgliali sono cellule fagocitiche appartenenti al sistema dei fagociti mononucleari e originate da cellule staminali ematopoietiche. La sua funzione è quella di proteggere dalle infezioni e dai danni e di rimuovere i prodotti di distruzione del tessuto nervoso. Le cellule microgliali sono caratterizzate da piccole dimensioni e corpi allungati. I loro brevi processi presentano sulla superficie rami secondari e terziari, che conferiscono alle cellule un aspetto “spinoso” (vedi Fig. 10.8). A differenza di altre cellule neurogliali, che hanno nuclei sferici, i nuclei dei microgliociti sono allungati, con cromatina compatta. La struttura descritta è caratteristica della microglia tipica (ramificata, a riposo) di un sistema nervoso centrale completamente formato. Ha una debole attività fagocitaria. Le microglia ramificate si trovano sia nella sostanza grigia che in quella bianca del sistema nervoso centrale. Una forma transitoria di microglia, la microglia ameboide, si trova nel cervello dei mammiferi in via di sviluppo. Le cellule microgliali ameboidi formano pieghe di filopodia e plasmalemma. Il loro citoplasma contiene numerosi fagolisosomi e corpi lamellari. Le cellule microgliali ameboidi sono caratterizzate da un'elevata attività degli enzimi lisosomiali. Le microglia ameboidi fagocitiche attive sono necessarie nel primo periodo postnatale, quando la barriera emato-encefalica non è ancora
è completamente sviluppato e le sostanze del sangue entrano facilmente nel sistema nervoso centrale. Si ritiene inoltre che aiuti a rimuovere i frammenti cellulari che compaiono a seguito della morte programmata dei neuroni in eccesso e dei loro processi. Si ritiene che, quando maturano, le cellule microgliali ameboidi si trasformino in microgliociti ramificati.
Oltre ai macrofagi gliali, ci sono cellule microgliali, classificate come “astrociti a riposo”. Questi ultimi sono capaci di proliferazione e differenziazione in astrociti.
Microglia reattiva appare dopo un infortunio in qualsiasi area del cervello. Le cellule microgliali si moltiplicano rapidamente e si attivano, il che si manifesta con la fagocitosi. In alcune malattie del sistema nervoso viene rilevata anche l'attività fagocitaria dei microgliociti (morbo di Alzheimer, encefalite autoimmune, ecc.). Un microgliocita attivato non ha processi di ramificazione, come una cellula a riposo, e non ha pseudopodi e filopodi, come le cellule microgliali ameboidi. Il citoplasma delle cellule microgliali reattive contiene corpi densi, inclusioni lipidiche e lisosomi.
Glia del sistema nervoso periferico(nevroglia periferica), a differenza della macroglia del sistema nervoso centrale, provengono dalla cresta neurale. La neuroglia periferica comprende neurolemmociti (cellule di Schwann) e gliociti gangliari (gliociti satelliti).
Neurolemmociti formano le guaine dei processi delle cellule nervose nelle fibre nervose del sistema nervoso periferico (vedi Fig. 10.9). Gliociti gangliari (gliociti ganglii) circondano i corpi cellulari dei neuroni nei gangli e partecipano al metabolismo dei neuroni.
10.4. FIBRE NERVOSE
I processi delle cellule nervose ricoperte da membrane sono chiamati fibre nervose (neurofibra). In base alla struttura delle guaine si distinguono le fibre nervose mielinizzate e non mielinizzate (Fig. 10.13, A, B). Il processo di una cellula nervosa in una fibra nervosa è chiamato cilindro assiale, o assone, poiché molto spesso (ad eccezione dei nervi sensoriali) le fibre nervose contengono assoni.
Nel sistema nervoso centrale, le guaine degli assoni e dei dendriti dei neuroni formano oligodendro-gliociti, e nel sistema nervoso periferico - neurolemmociti.
10.4.1. Fibre nervose non mielinizzate
Fibre nervose non mielinizzate (neurofibra amielinata) si trovano principalmente nel sistema nervoso autonomo. Nelle fibre nervose non mielinizzate, i processi delle cellule nervose sono immersi in depressioni sulla superficie dei neurolemmociti. Immerso nel corpo della cellula gliale

Riso. 10.13. La struttura delle fibre nervose a livello ottico-luce (A, B) e ultramicroscopico (a, b) (secondo T. N. Radostina, Yu. I. Afanasyev, L. S. Rumyantseva): A, UN- fibra mielinica; B, B- fibra non mielinizzata. 1 - cilindri assiali; 2 - strato di mielina; 3 - tessuto connettivo; 4 - tacca mielinica; 5 - nucleo dei neurolemmociti; 6 - intercettazione nodale; 7 - microtubuli; 8 - neurofilamenti; 9 - mitocondri; 10 - mesesone; 11 - membrana basale
Il processo neurale è limitato sia dal proprio plasmalemma che dallo stretto bordo del citoplasma del neurolemmocita. Nelle fibre nervose non mielinizzate degli organi interni, diversi cilindri assiali (10-20) appartenenti a diversi neuroni possono essere immersi nel citoplasma di un neurolemmocita. Spesso i cilindri assiali lasciano una fibra e passano in una fibra nervosa adiacente. La microscopia elettronica delle fibre nervose non mielinizzate mostra che quando i cilindri assiali vi si immergono,
i rolelemmociti del plasmalemma di quest'ultimo si piegano, avvolgono strettamente i cilindri assiali e, chiudendosi sopra di essi, formano pieghe profonde, sul fondo delle quali si trovano i singoli cilindri assiali. Le aree del plasmalemma del neurolemmocito, riunite nella zona della piega, formano una doppia membrana - Mesassone, sul quale sembra sospeso il cilindro assiale (vedi Fig. 10.13, B, b).
10.4.2. Fibre nervose mietoniche
Fibre nervose mielinizzate (neurofibra mielinata) presenti sia nel sistema nervoso centrale che periferico. Sono molto più spesse delle fibre nervose non mielinizzate. Il loro diametro della sezione trasversale varia da 2 a 20 micron. Anch'esse sono costituite da un cilindro assiale ricoperto da una guaina di neurolemmociti (cellule di Schwann), ma il diametro dei cilindri assiali di questo tipo di fibra è molto più spesso e la guaina è più complessa. Nella fibra mielinica formata, è consuetudine distinguere due strati della guaina: quello interno, più spesso, - strato di mielina (strato mielinico)(vedi Fig. 10.13, Aa) ed esterno, sottile, costituito da citoplasma, nuclei di neurolemmociti e neurolemma (neurolemma).
Lo strato di mielina contiene una quantità significativa di lipidi, quindi se trattato con acido osmico diventa marrone scuro. Nello strato mielinico si trovano periodicamente sottili linee luminose - incisioni sulla mielina (incisura myelini), o tacche Schmidt-Lanterman. A determinati intervalli (1-2 mm) sono visibili tratti di fibra privi dello strato mielinico - intercettazioni nodali (nodusruptionis myelini), o nodi di Ranvier.
Durante la formazione della fibra nervosa mielinizzata, il cilindro assiale non è semplicemente immerso nel citoplasma del neurolemmocita, ma è circondato da una membrana a strati a spirale formata avvolgendo il mesassone del neurolemmocita mentre ruota attorno al processo della cellula nervosa. Mentre ruota, il mesassone si allunga e si stratifica concentricamente sul cilindro assiale, formando attorno ad esso una densa zona stratificata - strato di mielina. Le micrografie elettroniche mostrano le principali linee dense e intraperiodiche. I primi sono formati dalla fusione delle superfici citoplasmatiche del plasmalemma del neurolemmocita (o oligodendrogliocita nel sistema nervoso centrale), i secondi dal contatto delle superfici extracellulari degli strati adiacenti del plasmalemma del neurolemmocita (Fig. 10.14 ). L'assenza di uno strato di mielina nell'area delle intercettazioni nodali è spiegata dal fatto che in questa sezione della fibra termina un neurolemmocita e ne inizia un altro. Il cilindro assiale in questo posto è parzialmente coperto da processi interdigitanti di neurolemmotsit. L'axolemma (guaina dell'assone) ha una densità elettronica significativa nell'area di intercettazione. La presenza di un gran numero di mitocondri in quest'area indica l'elevata attività metabolica dell'axolemma. L'axolemma dell'intercettazione possiede molti canali Na+ voltaggio-dipendenti necessari per la conduzione degli impulsi nervosi. Dovrebbe essere notato

È chiaro che la ramificazione assonale avviene anche nella zona delle intercettazioni.
Viene chiamata la lunghezza della fibra tra intercettazioni adiacenti segmento internodale. La lunghezza del segmento internodale, così come lo spessore dello strato mielinico, dipende dallo spessore del cilindro assiale. L'incisura mielinica è una sezione dello strato mielinico in cui i riccioli mesassoni giacciono sciolti l'uno rispetto all'altro, formando un tunnel a spirale che va dall'esterno verso l'interno e riempito con il citoplasma del neurolemmocita, cioè il luogo della dissezione della mielina. All'esterno del neurolemmocita è presente una membrana basale.
Fibre mielinizzate del sistema nervoso centrale differiscono in quanto in essi lo strato mielinico forma uno dei processi dell'oligodendrogliocita. I suoi restanti processi partecipano alla formazione dello strato mielinico di altre fibre mieliniche (ciascuna all'interno di un segmento internodale) (vedi Fig. 10.12). Le fibre mielinizzate del sistema nervoso centrale non hanno tacche mieliniche e non sono circondate da membrane basali. La mielina nel sistema nervoso centrale contiene proteine alcaline mieliniche e proteine proteolipide. Diverse malattie demielinizzanti del sistema nervoso centrale umano sono associate alla carenza o all'assenza di una o entrambe le proteine.
La velocità di trasmissione degli impulsi da parte delle fibre mielinizzate è maggiore di quella delle fibre non mielinizzate. Le fibre sottili povere di mielina e le fibre non mielinizzate conducono gli impulsi nervosi ad una velocità di 1-2 m/s, mentre le fibre mieliniche spesse conducono un impulso nervoso ad una velocità di 5-120 m/s.
In una fibra non mielinizzata l'onda di depolarizzazione della membrana viaggia lungo tutto l'assolemma senza interruzione, mentre in una fibra mielinizzata avviene solo nella zona di intercettazione, fornita dai canali del Na+. Pertanto, le fibre mielinizzate sono caratterizzate dalla conduzione saltatoria dell'eccitazione, cioè dal salto. Tra le intercettazioni passa una corrente elettrica la cui velocità è superiore al passaggio dell'onda di depolarizzazione lungo l'assolemma.
Riso. 10.14. Sviluppo e struttura della fibra mielinica (diagramma): UN- sezioni trasversali di stadi successivi di sviluppo della fibra mielinica (secondo Robertson); B- immagine tridimensionale della fibra formata (secondo M. H. Ross, L. J. Romrell). 1 - duplicazione della membrana dei neurolemmociti (mesassone); 2 - assone; 3 - tacca mielinica; 4 - contatti simili a dita del neurolemmocita nell'area di intercettazione; 5 - citoplasma dei neurolemmociti; 6 - mesassone ritorto a spirale; 7 - nucleo dei neurolemmociti
10.4.3. Risposta dei neuroni e delle loro fibre al danno
La sezione di una fibra nervosa provoca reazioni diverse nel corpo del neurone, nella sezione della fibra tra il corpo del neurone e il sito di sezione (segmento prossimale) e nella sezione situata distalmente al sito di lesione e non collegata al corpo del neurone (segmento distale). I cambiamenti nel corpo del neurone (perikaryon) sono espressi nel suo gonfiore, tigrolisi - dissoluzione di grumi di sostanza cromatofila e nel movimento del nucleo verso la periferia del corpo cellulare. I cambiamenti degenerativi nel segmento centrale sono limitati alla disintegrazione dello strato mielinico e del cilindro assiale vicino alla lesione. Nel segmento distale, lo strato mielinico e il cilindro assiale sono frammentati e i prodotti di degradazione vengono rimossi dai macrofagi, solitamente entro 1 settimana (Fig. 10.15).
La rigenerazione dipende dalla posizione della lesione. Sia nel sistema nervoso centrale che in quello periferico, i neuroni morti non vengono ripristinati. Di solito non si verifica una rigenerazione completa delle fibre nervose nel sistema nervoso centrale, ma le fibre nervose nei nervi periferici di solito si rigenerano bene. In questo caso i neurolemmociti del segmento periferico e del segmento centrale più vicini alla zona della lesione proliferano e si allineano in corde compatte. Il cono di crescita dell'assone si muove ad una velocità di 1-3 mm al giorno lungo la superficie dei neurolemmociti, staccando la membrana basale che ricopre le cellule. I neurolemmociti stimolano la crescita dell'assone, la direzione della sua crescita verso il bersaglio.
Se c'è un ostacolo alla crescita degli assoni del segmento centrale del nervo nelle corde dei neurolemmociti del segmento periferico (trauma esteso, infiammazione, presenza di una cicatrice), gli assoni del segmento centrale crescono in modo casuale e possono formare un groviglio chiamato neuroma da amputazione. Quando è irritato, si manifesta un forte dolore, che viene percepito come proveniente dalla zona inizialmente innervata, ad esempio come dolore in un arto amputato (dolore fantasma). La capacità delle fibre nervose di rigenerarsi preservando il pericario viene utilizzata in microchirurgia quando si suturano i processi distali e prossimali di un nervo danneggiato. Se ciò non è possibile, vengono utilizzate delle protesi (un tratto di vena), in cui vengono inserite le estremità del nervo danneggiato.
Le fibre nervose danneggiate del cervello e del midollo spinale non si rigenerano, ad eccezione degli assoni dei neuroni neurosecretori dell'ipotalamo. La rigenerazione delle fibre nel sistema nervoso centrale può essere indotta sperimentalmente trapiantandovi un nervo periferico. Forse la rigenerazione delle fibre nervose nel sistema nervoso centrale non avviene perché i gliociti senza membrana basale mancano dei fattori chemiotattici necessari per la conduzione degli assoni rigeneranti. Tuttavia, con lievi lesioni al sistema nervoso centrale, è possibile un ripristino parziale delle sue funzioni, a causa della plasticità del tessuto nervoso.

Riso. 10.15. Rigenerazione della fibra nervosa dopo la sezione (secondo R.V. Krstic): UN- fibra nervosa normale (nel corpo del neurone sono visibili una sostanza cromatofila e un nucleo al centro); B, V- fibra nervosa 2 settimane dopo il suo danno (la sostanza cromatofila nel corpo del neurone si riduce, il nucleo si sposta verso la periferia, la parte distale della fibra degenera, i prodotti di decadimento vengono fagocitati dai macrofagi); G - fibra nervosa 3 settimane dopo la transezione (la fibra muscolare si atrofizza, i neurolemmociti proliferano formando corde in cui è incorporato l'assone che cresce dalla parte centrale; aumenta la quantità di sostanza cromatofila nel pericario); D- fibra nervosa 3 mesi dopo la sua sezione (la struttura della fibra nervosa, del pericario e della fibra muscolare viene ripristinata); e- ridotta crescita degli assoni e formazione di cicatrici nel tessuto connettivo. 1 - cilindro assiale; 2 - perikaryon (corpo del neurone); 3 - frammentazione della mielina e formazione di goccioline di grasso; 4 - placca motore; 5 - neurolemmociti; 6 - microglia (macrofagi); 7 - mitosi delle cellule di Schwann e formazione di bande di Büngner; 8 - fibra muscolare; 9 - neuroma da amputazione; P - nodo di Ranvier
Le fibre nervose terminano nell'apparato terminale - terminazioni nervose (terminationis nervorum). Esistono tre gruppi di terminazioni nervose: apparati terminali che formano le sinapsi interneuronali e comunicano tra i neuroni; terminazioni effettrici (effettori), che trasmettono impulsi nervosi ai tessuti dell'organo funzionante; recettore (affettivo o sensibile).
10.5.1. Sinapsi
Sinapsi (sinapsi)- si tratta di contatti intercellulari specializzati progettati per trasmettere impulsi da un neurone all'altro o alle strutture muscolari e ghiandolari. Le sinapsi provvedono alla polarizzazione della trasmissione degli impulsi lungo una catena di neuroni, cioè determinano la direzione della trasmissione degli impulsi. Se stimoli un assone con una corrente elettrica, l'impulso andrà in entrambe le direzioni, ma l'impulso che va verso il corpo del neurone e i suoi dendriti non può essere trasmesso ad altri neuroni. Solo un impulso che raggiunge i terminali dell'assone può trasmettere l'eccitazione attraverso le sinapsi ad un altro neurone, muscolo o cellula ghiandolare. A seconda del metodo di trasmissione degli impulsi, le sinapsi possono essere chimico O elettrico(elettrotonico).

Riso. 10.16. Struttura delle sinapsi:
A - diagramma della citotopografia delle sinapsi; B - diagramma della struttura delle sinapsi: UN- tipo di freno; B- tipo eccitatorio; V- di tipo elettrico (senza bolle).
Sinapsi interneuronali
A seconda della localizzazione delle terminazioni dei rami terminali dell'assone del primo neurone, si distinguono le sinapsi assodendritiche, assospinose, assosomatiche e assoassonali (Fig. 10.16).
Le sinapsi chimiche trasmettono un impulso a un'altra cellula con l'aiuto di speciali sostanze biologicamente attive - neurotrasmettitori situati nelle vescicole sinaptiche (vedi Fig. 10.16, c, d). Il terminale dell'assone è presinaptico parte, e l'area del secondo neurone, o altra cellula innervata con cui è in contatto, - postsinaptico Parte.

Riso. 10.16. Continuazione
B - diagramma della struttura delle vescicole sinaptiche: UN- colinergico (leggero); B- adrenergici; V- purinergico; G- peptidergico (secondo L. D. Markina); D - micrografia elettronica di una sinapsi assodendritica (preparazione di I. G. Pavlova). 1 - sinapsi assosomatica; 2 - sinapsi assodendritiche; 3 - sinapsi assoassonale; 4 - dendriti; 5 - spina dendritica; 6 - assone; 7 - vescicole sinaptiche; 8 - membrana presinaptica; 9 - membrana postsinaptica; 10 - fessura sinaptica; 11 - sigilli postsinaptici
La parte presinaptica contiene vescicole sinaptiche, numerosi mitocondri e singoli neurofilamenti. La forma e il contenuto delle vescicole sinaptiche sono legati alla funzione della sinapsi. Ad esempio, nelle sinapsi in cui la trasmissione degli impulsi avviene con l'aiuto dell'acetilcolina (sinapsi colinergiche) sono presenti vescicole rotonde trasparenti con un diametro di 30-50 nm. Colinergiche sono le sinapsi simpatiche parasimpatiche e pregangliari, le sinapsi axomuscolari (vedi sotto) e alcune sinapsi del sistema nervoso centrale. Nelle sinapsi in cui viene utilizzato il neurotrasmettitore norepinefrina(sinapsi adrenergiche), sono presenti vescicole sinaptiche con un diametro di 50-90 nm con un nucleo denso di elettroni con un diametro di 15-25 nm. La norepinefrina è un mediatore delle sinapsi simpatiche postgangliari. Acetilcolina e norepinefrina sono i neurotrasmettitori più comuni, ma ce ne sono molti altri. Esistono neurotrasmettitori a basso peso molecolare, cioè con peso molecolare relativo piccolo (acetilcolina, norepinefrina, dopamina, glicina, acido gamma-aminobutirrico, serotonina, istamina, glutammato) e neuropeptidi: oppioidi (endorfine, encefaline), sostanza P, ecc. La dopamina, la glicina e l'acido gamma-aminobutirrico sono mediatori delle sinapsi inibitorie. Le endorfine e le encefaline prodotte nel cervello sono inibitori della percezione del dolore. Tuttavia, la maggior parte dei trasmettitori e, di conseguenza, le sinapsi sono eccitatorie. L'area di contatto sinaptico tra due neuroni è costituita da una membrana presinaptica, una fessura sinaptica e una membrana postsinaptica.
Membrana presinaptica- è il plasmalemma della cellula che trasmette l'impulso (axolemma). Contiene aree di ispessimento - zone attive in cui si verifica l'esocitosi del neurotrasmettitore. Le zone si trovano di fronte ai gruppi di recettori nella membrana postsinaptica. La membrana plasmatica nella zona attiva contiene canali Ca 2 + voltaggio-dipendenti. Quando la membrana è depolarizzata, i canali si aprono, favorendo l'esocitosi del neurotrasmettitore.
Fessura sinaptica tra le membrane pre e postsinaptica ha una larghezza di 20-30 nm. Le membrane sono saldamente attaccate tra loro nella regione sinaptica tramite filamenti che attraversano la fessura sinaptica.
Membrana postsinaptica- questa è una sezione del plasmalemma cellulare che contiene recettori dei neurotrasmettitori e canali ionici. Qui, le densità postsinaptiche spesse 20-70 nm si trovano sotto forma di una formazione omogenea densa di elettroni o di singoli corpi di forma rotonda. Le condensazioni sono costituite da una base granulare filamentosa che è integrata con il citoscheletro postsinaptico.
In generale, i processi nella sinapsi avvengono nel seguente ordine: 1) l'onda di depolarizzazione raggiunge la membrana presinaptica; 2) i canali del calcio si aprono e il Ca 2+ entra nel terminale; 3) l'ingresso di Ca 2+ nel terminale provoca l'esocitosi del neurotrasmettitore; in questo caso la membrana delle vescicole sinaptiche fa parte della membrana presinaptica e il trasmettitore entra nella fessura sinaptica; successivamente, le membrane delle vescicole sinaptiche, che divennero parte della membrana presinaptica, e parte della media

Riso. 10.17. Cambiamenti ciclici nelle vescicole sinaptiche nella sinapsi (secondo G. R. Nobak, N. L. Strominger, R. J. Demarest):
I - fibra nervosa; II - sinapsi; III - parte presinaptica. 1 - microtubuli;
2 - guaina mielinica; 3 - formazione di cisterne dalle quali si riformano le vescicole sinaptiche; 4 - formazione di nuove membrane di vescicole sinaptiche mediante pinocitosi (endocitosi) di porzioni del neurotrasmettitore; 5 - fessura sinaptica; 6 - membrana postsinaptica; 7 - fusione della membrana della vescicola sinaptica con il plasmalemma e rilascio del neurotrasmettitore per esocitosi nella fessura sinaptica; 8 - vescicole sinaptiche; 9 - mitocondri
il toro subisce endocitosi e si verifica il ricircolo delle vescicole sinaptiche (Fig. 10.17) e parte delle membrane e del neurotrasmettitore entra nel pericario utilizzando il trasporto retrogrado e viene distrutto dai lisosomi; 4) la molecola del neurotrasmettitore si lega ai siti recettoriali sulla membrana postsinaptica, provocando 5) cambiamenti molecolari nella membrana postsinaptica, che portano a 6) l'apertura di canali ionici e 7) la creazione di potenziali postsinaptici che causano reazioni di eccitazione o inibizione; 8) la rimozione del neurotrasmettitore dalla fessura avviene a causa della sua scissione da parte di un enzima e rimozione mediante cattura da parte di un trasportatore specifico.
Le sinapsi elettriche o elettrotoniche sono relativamente rare nel sistema nervoso dei mammiferi. Nell'area di tali sinapsi, i citoplasmi dei neuroni vicini sono collegati da giunzioni gap (contatti), garantendo il passaggio degli ioni da una cellula all'altra,

Riso. 10.18. Struttura ultramicroscopica della giunzione neuromuscolare (schema): 1 - citoplasma del neurolemmocito; 2 - nucleo dei neurolemmociti; 3 - plasmalemma del neurolemmocita; 4 - cilindro assiale della fibra nervosa; 5 - axolemma; 6 - membrana postsinaptica (sarcolemma); 7 - mitocondri nell'assoplasma; 8 - fessura sinaptica; 9 - mitocondri nel sarcoplasma della fibra muscolare; 10 - vescicole presinaptiche; 11 - membrana presinaptica (axolemma); 12 - sarcolemma; 13 - nucleo della fibra muscolare; 14 - miofibrilla
e quindi l'interazione elettrica di queste cellule. Queste sinapsi aiutano a sincronizzare l'attività.
Le strutture sinaptiche sono altamente sensibili all'azione di fattori tossici e di sostanze tossiche psicotrope. I disturbi nella trasmissione degli impulsi nervosi nell'area delle sinapsi (acquisiti o determinati geneticamente) sono alla base dello sviluppo di una serie di malattie del sistema nervoso umano.
10.5.2. Terminazioni nervose effettrici
Le terminazioni nervose effettrici sono di due tipi: motorie e secretorie.
Terminazioni nervose motorie- questi sono i dispositivi terminali degli assoni delle cellule motorie del sistema nervoso somatico o autonomo. Con la loro partecipazione, l'impulso nervoso viene trasmesso ai tessuti degli organi funzionanti. La terminazione motoria dei muscoli striati è chiamata giunzione neuromuscolare o sinapsi. (sinapsi neuromuscolare). La giunzione neuromuscolare è costituita dalla ramificazione terminale del cilindro assiale della fibra nervosa e da un tratto specializzato della fibra muscolare (Fig. 10.18). La fibra nervosa mielinizzata, avvicinandosi alla fibra muscolare, perde uno strato di mielina e forma un terminale neuromuscolare specializzato.

Riso. 10.19. Terminazioni nervose motorie nel tessuto muscolare liscio: 1 - corpo (pericario) di un neurone multipolare; 2 - dendriti; 3 - assone; 4 - ispessimenti con vescicole sinaptiche; 5 - vescicole sinaptiche; 6 - miociti lisci
zione. I neurolemmociti sono appiattiti, la loro membrana basale continua nella membrana basale della fibra muscolare. Il plasmalemma dei rami terminali dell'assone e il sarcolemma della fibra muscolare sono separati da una fessura sinaptica larga circa 50 nm. La fessura sinaptica è riempita da una sostanza amorfa ricca di glicoproteine. Il sarcolemma della fibra muscolare forma numerose pieghe che formano fessure sinaptiche secondarie della giunzione neuromuscolare. In questa zona la fibra muscolare non presenta le tipiche striature trasversali ed è caratterizzata da un'abbondanza di mitocondri, un ammasso di nuclei rotondi o leggermente ovali. Il sarcoplasma con mitocondri e nuclei costituisce insieme la parte postsinaptica della sinapsi.
I rami terminali della fibra nervosa alla giunzione neuromuscolare sono caratterizzati da un'abbondanza di mitocondri e da numerose vescicole presinaptiche contenenti un trasmettitore caratteristico di questo tipo di terminazioni - acetilcolina. Quando eccitata, l'acetilcolina entra nella fessura sinaptica attraverso la membrana presinaptica fino ai recettori colinergici della membrana postsinaptica (muscolare), provocandone l'eccitazione (onda di depolarizzazione).
La membrana postsinaptica della sinapsi neuromuscolare contiene l'enzima acetilcolinesterasi, che distrugge il trasmettitore e quindi ne limita la durata d'azione. I disturbi delle giunzioni neuromuscolari causano lo sviluppo di malattie incurabili miastenia grave, caratterizzata da progressiva debolezza muscolare che spesso termina con la paralisi dei muscoli respiratori (muscoli intercostali e diaframma). Con questa malattia circolano nel sangue anticorpi contro i recettori dell'acetilcolina del sarcolemma. Questi anticorpi si legano ai recettori colinergici della membrana postsinaptica e li inattivano, interrompendo l'interazione neuromuscolare.
Le terminazioni nervose motorie nel tessuto muscolare liscio sono ispessimenti a forma di perla fibra nervosa che corre tra i miociti lisci (Fig. 10.19). Gli ispessimenti contengono vescicole presinaptiche adrenergiche o colinergiche. I neurolemmociti nell'area di questi ispessimenti sono spesso assenti.
Hanno una struttura simile terminazioni nervose secretrici(neuroghiandolare - terminazione neuroghiandolare). Sono ispessimenti terminali di terminali o ispessimenti lungo la fibra nervosa, contenenti vescicole presinaptiche, prevalentemente colinergiche.
10.5.3. Terminazioni nervose recettrici
Queste terminazioni nervose - recettori - sono sparse in tutto il corpo e percepiscono varie irritazioni sia dall'ambiente esterno che dagli organi interni. Di conseguenza, si distinguono due grandi gruppi di recettori: esterocettori E interorecettori. Gli esterocettori (esterni) comprendono i recettori uditivi, visivi, olfattivi, gustativi e tattili. Gli interorecettori (interni) includono viscerorecettori (segnalazione sullo stato degli organi interni) e vestiboloproprio-recettori (recettori del sistema muscolo-scheletrico). A seconda della specificità dell'irritazione percepita da un dato tipo di recettore, tutte le terminazioni sensoriali sono suddivise in meccanocettori, barocettori, chemocettori, termorecettori e così via.
Secondo la loro struttura, le terminazioni sensibili sono divise in terminazioni nervose libere (terminatio nervi libera), che sono dendriti ramificati sottili senza guaina gliale, e non libero, contenente tutti i componenti della fibra nervosa, cioè i rami del cilindro assiale e le cellule gliali. Le terminazioni non libere, inoltre, possono essere ricoperte da una capsula di tessuto connettivo e quindi vengono chiamate incapsulate (coipusculum nervososum capsulatum). Le terminazioni nervose non libere che non hanno una capsula di tessuto connettivo sono chiamate non incapsulate (corpuscolo nervoso non capsulato)(Fig. 10.20).
Le terminazioni nervose libere tipicamente percepiscono il freddo, il caldo e il dolore. Tali terminazioni sono caratteristiche dell'epitelio. In questo caso, le fibre nervose mielinizzate si avvicinano allo strato epiteliale, perdono la mielina e i cilindri assiali penetrano nell'epitelio e lì si disintegrano tra le cellule in sottili rami terminali.
I recettori nel tessuto connettivo sono molto diversi. La stragrande maggioranza di essi rappresenta vari gradi di complessità della ramificazione del cilindro assiale. La composizione di tali apparati terminali, di regola, comprende neurolemmociti, che accompagnano tutti i rami delle fibre (si tratta di recettori non liberi e non incapsulati).
I recettori del tessuto connettivo incapsulati, con tutta la loro diversità, sono sempre costituiti da cilindri assiali ramificati e cellule gliali. All'esterno tali recettori sono ricoperti da una capsula di tessuto connettivo.

Riso. 10.20. Terminazioni nervose recettoriali (secondo R.V. Krstic, con modifiche): UN- terminazioni nervose libere (dolore); B- Il corpo di Meissner (tatto); V- Fiaschetta Krause (fredda); G- Corpo Vater-Pacini (pressione); D- Corpuscolo di Ruffini (calore)
Le terminazioni incapsulate sensibili includono i corpuscoli tattili (corpuscolo tatto)- I corpuscoli di Meissner. Si tratta di strutture di forma ovoidale, che misurano 50-150X60 micron. Si trovano nella parte superiore delle papille del tessuto connettivo della pelle. I corpuscoli tattili sono costituiti da neurolemmociti modificati: cellule tattili situate perpendicolari all'asse lungo del corpuscolo. Le parti delle cellule tattili contenenti nuclei si trovano alla periferia, e le parti appiattite rivolte verso il centro formano processi lamellari che si interdigitano con i processi del lato opposto (Fig. 10.21). Il corpo è circondato da una capsula sottile. La fibra nervosa mielinizzata entra dal basso nella base del corpuscolo, perde lo strato di mielina e forma rami che si snodano tra le cellule tattili. Microfibrille e fibre di collagene si legano tattilmente

Riso. 10.21. Corpo tattile nel tessuto connettivo della pelle (micrografia). Impregnazione con nitrato d'argento
cellule con una capsula e la capsula con lo strato basale dell'epidermide, in modo che qualsiasi spostamento dell'epidermide venga trasmesso al corpo tattile.
I corpi lamellari sono molto diffusi nell'uomo (corpuscolo lamelloso- Corpuscoli di Vater-Pacini). Le loro dimensioni sono 0,5X1-2 mm. Al centro di tale corpo c'è un bulbo interno, o pallone. (bulbo interno), formato da lemmociti modificati (Fig. 10.22). La fibra nervosa mielinica sensibile perde il suo nuovo strato di mielina in prossimità del corpo lamellare, penetra nel bulbo interno e si ramifica. All'esterno, il corpo è circondato da una capsula stratificata costituita da fibroblasti e fibre orientate a spirale. Gli spazi pieni di liquido tra le piastre contengono microfibrille di collagene. La pressione sulla capsula viene trasmessa attraverso gli spazi pieni di liquido tra le piastre al bulbo interno e viene ricevuta dalle fibre non mielinizzate nel bulbo interno. I corpi lamellari percepiscono la pressione e la vibrazione. Sono presenti negli strati profondi del derma (soprattutto nella pelle delle dita), nel mesentere e negli organi interni.
Le terminazioni nervose incapsulate comprendono anche i recettori muscolari e tendinei: i fusi neuromuscolari (fusus neuromuscolare) e fusi neurotendinei (fuso neurotendineo)(Fig. 10.23).
I fusi neuromuscolari sono organi sensoriali nel muscolo scheletrico che funzionano come recettori di stiramento. Il fuso è costituito da numerose fibre muscolari striate racchiuse in una capsula estensibile di tessuto connettivo - intrafusale fibre Vengono chiamate le restanti fibre muscolari che si trovano all'esterno della capsula extrafusale. La capsula ha una struttura a strati. Distingue tra strati esterni e interni. C'è uno spazio pieno di liquido tra la capsula e le fibre intrafusali.

Riso. 10.22. Struttura ultramicroscopica delle terminazioni nervose incapsulate: UN- corpo lamellare di Vater-Pacini: 1 - capsula stratificata: 2 - bulbo interno: 3 - dendrite di cellula nervosa sensitiva; 4 - fibre di collagene a spirale; 5 - fibrociti; 6 - cellule sensoriali secondarie con ciglia; 7 - contatti sinaptici degli assoni delle cellule sensoriali secondarie con i dendriti della cellula nervosa sensibile (secondo A. A. Otelin, V. R. Mashansky, A. S. Mirkin); B- Corpuscolo tattile di Meissner: 1 - capsula; 2 - celle speciali; 3 - terminali nervosi; 4 - fibra nervosa mielinica; 5 - fibrille di supporto (di supporto); 6 - epitelio (secondo R.V. Krstic, con modifiche)
La parte recettore della fibra muscolare intrafusale è la parte centrale, non contrattile. Esistono due tipi di fibre intrafusali: fibre con una borsa nucleare (bursa nuclearis) E fibre con una catena nucleare (vinculum nucleare). Il fuso umano contiene da 1 a 3 fibre con una borsa nucleare. Quello centrale esteso contiene molti nuclei. Le fibre con una catena nucleare nel fuso possono essere da 3 a 7. Sono due volte più sottili e lunghe la metà delle fibre con una sacca nucleare, e i nuclei in esse si trovano in una catena in tutta la regione del recettore. Due tipi di fibre afferenti si avvicinano alle fibre muscolari intrafusali: primarie e secondarie. Le fibre primarie con un diametro di 17 micron formano terminazioni a forma di spirale - terminazioni ad anello-spirale (terminatio nervi annu-lospiralis)- sia su fibre con sacco nucleare che su fibre con catena nucleare. Le fibre secondarie con un diametro di 8 μm innervano le fibre con la catena nucleare. Su entrambi i lati dell'estremità a spirale dell'anello formano terminazioni a forma di uva (terminazione nervosa racemosa).
Quando il muscolo si rilassa (o si allunga), aumenta anche la lunghezza delle fibre intrafusali, che viene registrata dai recettori. Anello-spirale

Riso. 10.23. Struttura del fuso neuromuscolare (schema):
UN- innervazione motoria delle fibre muscolari intrafusali ed extrafusali (secondo A. N. Studitsky); B- terminazioni nervose afferenti a spirale attorno alle fibre muscolari intrafusali nell'area delle borse nucleari (secondo R.V. Krstic, con modifiche). 1 - terminazioni effettrici neuromuscolari delle fibre muscolari extrafusali; 2 - placche motorie delle fibre muscolari intrafusali; 3 - capsula del tessuto connettivo; 4 - borsa nucleare; 5 - terminazioni nervose sensibili ad anello-spirale attorno alle sacche nucleari; 6 - fibre muscolari scheletriche; 7 - nervo
le terminazioni reagiscono ai cambiamenti nella lunghezza della fibra muscolare e alla velocità di questo cambiamento, le terminazioni a forma di acino d'uva rispondono solo ai cambiamenti di lunghezza. Con un allungamento improvviso, un forte segnale viene inviato dalle terminazioni della spirale dell'anello al midollo spinale, provocando una forte contrazione del muscolo da cui proviene il segnale: un riflesso di stiramento dinamico. Con lo stiramento lento e a lungo termine della fibra, si verifica un segnale di trazione statico, trasmesso sia dai recettori a spirale ad anello che a forma di acino d'uva. Questo segnale può mantenere il muscolo in uno stato di contrazione per diverse ore.

Riso. 10.24. Arco riflesso semplice (schema secondo V. G. Eliseev, Yu. I. Afanasyev, E. F. Kotovsky):
1 - cellula nervosa sensibile; 2 - dendrite di una cellula sensibile; 3 - recettore nella pelle; 4 - plasmalemma del neurolemmocito; 5 - nuclei di neurolemmociti; 6 - strato di mielina; 7 - intercettazione nodale della fibra nervosa; 8 - cilindro assiale; 9 - tacca mielinica; 10 - assone di una cellula sensibile; 11 - cellula motoria (motoneurone); 12 - dendriti della cellula motoria; 13 - assone della cellula motoria; 14 - fibre mieliniche; 15 - effettore sul muscolo; 16 - nodo sensibile; 17 - ramo dorsale del nervo spinale; 18 - radice posteriore; 19 - corno posteriore; 20 - clacson anteriore; 21 - radice anteriore; 22 - ramo ventrale del nervo spinale
Le fibre intrafusali hanno anche innervazione efferente. Ad essi si avvicinano sottili fibre motorie, che terminano in sinapsi asmuscolari alle estremità della fibra muscolare. Provocando la contrazione delle sezioni terminali della fibra intrafusale, aumentano lo stiramento della sua parte recettrice centrale, aumentando la risposta recettoriale.
Fusi neurotendinei solitamente si trova alla giunzione tra muscolo e tendine. Fasci di fibre collagene tendinee associati a 10-15 fibre muscolari sono circondati da una capsula di tessuto connettivo. Una fibra mielinica spessa (16 µm di diametro) si avvicina al fuso nervo-tendineo, che perde mielina e forma terminali che si ramificano tra i fasci di fibre collagene del tendine. Il segnale proveniente dai fusi neurotendinei, causato dalla tensione muscolare, eccita i neuroni inibitori del midollo spinale. Questi ultimi inibiscono i corrispondenti motoneuroni, prevenendo l'eccessivo allungamento muscolare.
10.6. IL CONCETTO DI ARCO RIFLETTORE
Il tessuto nervoso fa parte del sistema nervoso, che funziona secondo il principio riflesso, il cui substrato morfologico è l'arco riflesso. Un arco riflesso è una catena di neuroni collegati tra loro da sinapsi e che garantisce la conduzione di un impulso nervoso dal recettore di un neurone sensibile all'effettore che termina nell'organo funzionante.
L'arco riflesso più semplice è costituito da due neuroni: sensibile e motorio (Fig. 10.24). Nella stragrande maggioranza dei casi, i neuroni intercalari o associativi sono compresi tra i neuroni sensoriali e quelli motori. Negli animali superiori, gli archi riflessi sono solitamente costituiti da molti neuroni e hanno una struttura molto più complessa rispetto alla figura mostrata. Connessioni neurali specifiche verranno esaminate utilizzando l'esempio della corteccia cerebrale e del cervelletto.
Domande di controllo
1. Fonti di sviluppo e classificazione dei neuroni, organizzazione ultrastrutturale.
2. Differenze cellulari del tessuto nervoso.
3. Neuroglia: classificazione, topografia del sistema nervoso, funzioni.
4. Sinapsi: struttura, funzioni, classificazione.
5. Archi riflessi come base morfologica dell'attività del tessuto nervoso.
Istologia, embriologia, citologia: libro di testo / Yu. I. Afanasyev, N. A. Yurina, E. F. Kotovsky, ecc. - 6a edizione, rivista. e aggiuntivi - 2012. - 800 p. : malato.
Siamo spesso nervosi, filtriamo costantemente le informazioni in arrivo, reagiamo al mondo che ci circonda e cerchiamo di ascoltare il nostro stesso corpo, e cellule straordinarie ci aiutano in tutto questo. Sono il risultato di una lunga evoluzione, il risultato del lavoro della natura durante tutto lo sviluppo degli organismi sulla Terra.
Non possiamo dire che il nostro sistema di percezione, analisi e risposta sia ideale. Ma siamo arrivati molto lontano dagli animali. Comprendere come funziona un sistema così complesso è molto importante non solo per gli specialisti: biologi e medici. Anche una persona di un'altra professione potrebbe essere interessata a questo.
Le informazioni contenute in questo articolo sono a disposizione di tutti e possono essere utili non solo come conoscenza, perché comprendere il proprio corpo è la chiave per comprendere se stessi.
Di cosa è responsabile?
Il tessuto nervoso umano si distingue per una diversità strutturale e funzionale unica dei neuroni e per la specificità delle loro interazioni. Dopotutto, il nostro cervello è un sistema molto complesso. E per controllare il nostro comportamento, le nostre emozioni e il nostro pensiero, abbiamo bisogno di una rete molto complessa.
Il tessuto nervoso, la cui struttura e funzioni sono determinate da un insieme di neuroni - cellule con processi - e determinano il normale funzionamento del corpo, in primo luogo, garantisce l'attività coordinata di tutti i sistemi di organi. In secondo luogo, collega il corpo con l'ambiente esterno e fornisce reazioni adattative ai suoi cambiamenti. In terzo luogo, controlla il metabolismo in condizioni mutevoli. Tutti i tipi di tessuto nervoso sono una componente materiale della psiche: sistemi di segnalazione - parola e pensiero, caratteristiche comportamentali nella società. Alcuni scienziati hanno ipotizzato che l'uomo abbia sviluppato notevolmente la sua mente, per la quale ha dovuto “sacrificare” molte abilità animali. Ad esempio, non abbiamo la vista e l’udito acuti di cui possono vantarsi gli animali.
Il tessuto nervoso, la cui struttura e funzione si basa sulla trasmissione elettrica e chimica, ha effetti chiaramente localizzati. A differenza del sistema umorale, questo sistema agisce istantaneamente.
Molti piccoli trasmettitori
Le cellule del tessuto nervoso - i neuroni - sono le unità strutturali e funzionali del sistema nervoso. La cellula neuronale è caratterizzata da una struttura complessa e da una maggiore specializzazione funzionale. La struttura di un neurone è costituita da un corpo eucariotico (soma), il cui diametro è di 3-100 micron, e da processi. Il soma di un neurone contiene un nucleo e un nucleolo con un apparato biosintetico che forma enzimi e sostanze inerenti alle funzioni specializzate dei neuroni. Questi sono corpi di Nissl: cisterne appiattite strettamente adiacenti del reticolo endoplasmatico ruvido, nonché un apparato di Golgi sviluppato.

Le funzioni di una cellula nervosa possono essere svolte continuamente grazie all'abbondanza di "stazioni energetiche" nel corpo che producono ATP - condrasomi. Il citoscheletro, rappresentato da neurofilamenti e microtubuli, svolge un ruolo di supporto. Nel processo di perdita delle strutture della membrana, viene sintetizzato il pigmento lipofuscina, la cui quantità aumenta con l'aumentare dell'età del neurone. Il pigmento melatonina si forma nei neuroni staminali. Il nucleolo è costituito da proteine e RNA, il nucleo del DNA. L'ontogenesi del nucleolo e dei basofili è determinata dalle reazioni comportamentali primarie delle persone, poiché dipendono dall'attività e dalla frequenza dei contatti. Il tessuto nervoso si riferisce all'unità strutturale di base, il neurone, sebbene esistano altri tipi di tessuti di supporto.
Caratteristiche della struttura delle cellule nervose
Il nucleo a doppia membrana dei neuroni è dotato di pori attraverso i quali penetrano e vengono eliminate le sostanze di scarto. Grazie all'apparato genetico avviene la differenziazione, che determina la configurazione e la frequenza delle interazioni. Un'altra funzione del nucleo è quella di regolare la sintesi proteica. Le cellule nervose mature non possono dividersi per mitosi e i prodotti di sintesi attivi geneticamente determinati di ciascun neurone devono garantire il funzionamento e l’omeostasi durante l’intero ciclo di vita. La sostituzione delle parti danneggiate e perse può avvenire solo a livello intracellulare. Ma ci sono anche delle eccezioni. Nell'epitelio alcuni gangli animali sono capaci di dividersi.

Le cellule del tessuto nervoso si distinguono visivamente per una varietà di dimensioni e forme. I neuroni hanno contorni irregolari a causa dei loro processi, spesso numerosi e troppo cresciuti. Questi sono conduttori viventi di segnali elettrici attraverso i quali si formano archi riflessi. Il tessuto nervoso, la cui struttura e funzioni dipendono da cellule altamente differenziate il cui ruolo è quello di percepire le informazioni sensoriali, codificarle attraverso impulsi elettrici e trasmetterle ad altre cellule differenziate, è in grado di fornire una risposta. È quasi istantaneo. Ma alcune sostanze, incluso l'alcol, lo rallentano notevolmente.
A proposito di assoni
Tutti i tipi di tessuto nervoso funzionano con la partecipazione diretta di processi dendritici e assoni. Axon è tradotto dal greco come "asse". Questo è un processo allungato che conduce l'eccitazione dal corpo ai processi di altri neuroni. Le punte dell'assone sono molto ramificate, ciascuna è in grado di interagire con 5000 neuroni e formare fino a 10mila contatti.
Il luogo del soma da cui si diramano gli assoni è chiamato collinetta dell'assone. Ciò che ha in comune con l'assone è che mancano di reticolo endoplasmatico ruvido, RNA e un complesso enzimatico.
Un po' di dendriti
Il nome di questa cella significa "albero". Come i rami, dal soma crescono processi corti e fortemente ramificati. Ricevono segnali e fungono da luoghi in cui si verificano le sinapsi. I dendriti, con l'aiuto di processi laterali - spine - aumentano la superficie e, di conseguenza, i contatti. I dendriti senza rivestimento, ma circondati dagli assoni, sono di natura lipidica e il loro effetto è simile alle proprietà isolanti del rivestimento di plastica o gomma dei cavi elettrici. Il punto di generazione dell'eccitazione - la collinetta dell'assone - appare nel punto in cui l'assone si allontana dal soma nella zona trigger.
La sostanza bianca dei tratti ascendenti e discendenti nel midollo spinale e nel cervello è formata da assoni attraverso i quali vengono trasmessi gli impulsi nervosi, svolgendo una funzione conduttrice: la trasmissione di un impulso nervoso. I segnali elettrici vengono trasmessi a varie parti del cervello e del midollo spinale, comunicando tra loro. In questo caso, gli organi esecutivi possono connettersi con i recettori. La materia grigia forma la corteccia cerebrale. Nel canale spinale ci sono centri di riflessi innati (starnuti, tosse) e centri vegetativi di attività riflessa dello stomaco, minzione e defecazione. Gli interneuroni, i corpi motori e i dendriti svolgono una funzione riflessa, eseguendo reazioni motorie.

Le caratteristiche del tessuto nervoso sono determinate dal numero di processi. I neuroni sono unipolari, pseudounipolari, bipolari. Il tessuto nervoso umano non contiene unipolare con uno. Nel multipolare c'è abbondanza di tronchi dendritici. Questa ramificazione non influisce in alcun modo sulla velocità del segnale.
Celle diverse: compiti diversi
Le funzioni di una cellula nervosa sono eseguite da diversi gruppi di neuroni. In base alla loro specializzazione, l'arco riflesso è suddiviso in neuroni afferenti o sensoriali che conducono gli impulsi dagli organi e dalla pelle al cervello.
I neuroni intercalari, o neuroni associativi, sono un gruppo di neuroni di commutazione o di connessione che analizzano e prendono decisioni, eseguendo le funzioni di una cellula nervosa.
I neuroni efferenti, o neuroni sensoriali, trasportano informazioni sulle sensazioni: impulsi dalla pelle e dagli organi interni al cervello.
I neuroni efferenti, effettori o motori, conducono impulsi - "comandi" dal cervello e dal midollo spinale a tutti gli organi funzionanti.
La particolarità dei tessuti nervosi è che i neuroni svolgono un lavoro complesso e prezioso nel corpo, quindi il lavoro primitivo quotidiano - fornire nutrimento, rimuovere i prodotti di decomposizione, la funzione protettiva spetta alle cellule neurogliali ausiliarie o al supporto delle cellule di Schwann.
Il processo di formazione delle cellule nervose
Nelle cellule del tubo neurale e della placca gangliare avviene la differenziazione, che determina le caratteristiche dei tessuti nervosi in due direzioni: quelle grandi diventano neuroblasti e neurociti. Le piccole cellule (spongioblasti) non si ingrandiscono e diventano gliociti. Il tessuto nervoso, i cui tipi di tessuti sono composti da neuroni, è costituito da tessuti primari e ausiliari. Le cellule di supporto (“gliociti”) hanno una struttura e una funzione speciali.

Quello centrale è rappresentato dai seguenti tipi di gliociti: ependimociti, astrociti, oligodendrociti; periferici - gliociti gangliari, gliociti terminali e neurolemmociti - cellule di Schwann. Gli ependimociti rivestono le cavità dei ventricoli del cervello e del canale spinale e secernono liquido cerebrospinale. Tipi di tessuto nervoso: gli astrociti a forma di stella formano tessuti di sostanza grigia e bianca. Le proprietà del tessuto nervoso - gli astrociti e la loro membrana gliale contribuiscono alla creazione di una barriera emato-encefalica: un confine strutturale-funzionale passa tra il tessuto connettivo liquido e quello nervoso.
Evoluzione del tessuto
La proprietà principale di un organismo vivente è l'irritabilità o la sensibilità. Il tipo di tessuto nervoso è determinato dalla posizione filogenetica dell'animale ed è caratterizzato da un'ampia variabilità, diventando più complesso nel processo di evoluzione. Tutti gli organismi richiedono determinati parametri di coordinazione e regolazione interna, una corretta interazione tra stimolo per l'omeostasi e stato fisiologico. Il tessuto nervoso degli animali, soprattutto quelli multicellulari, la cui struttura e funzioni hanno subito aromorfosi, contribuisce alla sopravvivenza nella lotta per l'esistenza. Negli idroidi primitivi, è rappresentato da cellule nervose stellate sparse in tutto il corpo e collegate da sottili processi intrecciati tra loro. Questo tipo di tessuto nervoso è chiamato diffuso.
Il sistema nervoso dei nematodi piatti e ascaridi è stelo, di tipo scaleno (ortogonale) costituito da gangli cerebrali accoppiati - gruppi di cellule nervose e tronchi longitudinali che si estendono da essi (connettivi), interconnessi da corde-commissure trasversali. Negli anelli, dal ganglio perifaringeo, collegato da corde, si diparte la catena nervosa addominale, in ciascun segmento della quale sono presenti due gangli nervosi vicini collegati da fibre nervose. In alcuni animali dal corpo molle, i gangli nervosi sono concentrati per formare il cervello. Gli istinti e l'orientamento spaziale negli artropodi sono determinati dalla cefalizzazione dei gangli del cervello accoppiato, dall'anello nervoso perifaringeo e dal cordone nervoso ventrale.

Nei cordati, il tessuto nervoso, i cui tipi di tessuti sono fortemente espressi, è complesso, ma tale struttura è evolutivamente giustificata. Si presentano diversi strati che si trovano sul lato dorsale del corpo sotto forma di tubo neurale, la cavità è il neurocele. Nei vertebrati si differenzia in cervello e midollo spinale. Man mano che si forma il cervello, si formano rigonfiamenti all'estremità anteriore del tubo. Se negli organismi multicellulari inferiori il sistema nervoso svolge un ruolo puramente di collegamento, negli animali altamente organizzati immagazzina informazioni, le recupera quando necessario e garantisce anche l'elaborazione e l'integrazione.
Nei mammiferi, questi rigonfiamenti cerebrali danno origine alle parti principali del cervello. E il resto del tubo forma il midollo spinale. Il tessuto nervoso, la cui struttura e funzioni sono uniche nei mammiferi superiori, ha subito cambiamenti significativi. Questo è il progressivo sviluppo della corteccia cerebrale e di tutte le parti che determinano il complesso adattamento alle condizioni ambientali e la regolazione dell'omeostasi.
Centro e periferia
Le parti del sistema nervoso sono classificate in base alla loro struttura funzionale e anatomica. La struttura anatomica è simile alla toponomastica, dove si distinguono il sistema nervoso centrale e quello periferico. comprende il cervello e il midollo spinale, mentre quello periferico è rappresentato da nervi, nodi e terminazioni. I nervi sono rappresentati da gruppi di processi esterni al sistema nervoso centrale, ricoperti da una comune guaina mielinica, e conducono segnali elettrici. I dendriti dei neuroni sensoriali formano i nervi sensoriali, gli assoni formano i nervi motori.
La combinazione di processi lunghi e brevi forma nervi misti. Accumulandosi e concentrandosi, i corpi dei neuroni formano nodi che si estendono oltre il sistema nervoso centrale. Le terminazioni nervose si dividono in recettore ed effettore. I dendriti, attraverso i rami terminali, convertono gli stimoli in segnali elettrici. E le terminazioni efferenti degli assoni si trovano negli organi funzionanti, nelle fibre muscolari e nelle ghiandole. La classificazione per funzionalità implica la divisione del sistema nervoso in somatico e autonomo.
Alcune cose le controlliamo, altre non possiamo controllarle.
Le proprietà del tessuto nervoso spiegano il fatto che obbedisce alla volontà di una persona, innervando il lavoro del sistema di supporto. I centri motori si trovano nella corteccia cerebrale. Autonomo, chiamato anche vegetativo, non dipende dalla volontà di una persona. In base alle proprie richieste, è impossibile accelerare o rallentare il battito cardiaco o la motilità intestinale. Poiché la sede dei centri autonomi è l'ipotalamo, il sistema nervoso autonomo controlla il funzionamento del cuore e dei vasi sanguigni, dell'apparato endocrino e degli organi addominali.

Il tessuto nervoso, di cui potete vedere la foto sopra, forma le divisioni simpatica e parasimpatica, che permettono loro di agire come antagonisti, producendo un effetto reciprocamente opposto. L'eccitazione in un organo provoca processi di inibizione in un altro. Ad esempio, i neuroni simpatici provocano forti e frequenti contrazioni delle camere cardiache, vasocostrizione e aumenti della pressione sanguigna, poiché viene rilasciata la norepinefrina. L'attività parasimpatica, rilasciando acetilcolina, aiuta ad indebolire il ritmo cardiaco, ad aumentare il lume delle arterie e ad abbassare la pressione sanguigna. Il bilanciamento di questi gruppi di mediatori normalizza il ritmo cardiaco.
Il sistema nervoso simpatico funziona durante i periodi di intensa tensione come paura o stress. I segnali sorgono nell'area delle vertebre toraciche e lombari. Il sistema parasimpatico si attiva durante il riposo e la digestione del cibo, durante il sonno. I corpi cellulari dei neuroni si trovano nel tronco e nell'osso sacro.
Studiando più in dettaglio le caratteristiche delle cellule di Purkinje, che sono a forma di pera con molti dendriti ramificati, si può vedere come avviene la trasmissione degli impulsi e rivelare il meccanismo delle fasi successive del processo.