Физиология процессов дыхания. Физиология дыхания у человека
Кислород, являющийся конечным акцептором электронов в дыхательной цепи, необходимо доставить в организме к каждой клетке. У животных выработались системы внешнего дыхания, функцией которых и является газообмен. В курсе зоологии получены исчерпывающие сведения на этот счет.
Дыхание – это совокупность процессов и механизмов, обеспечивающих потребление кислорода и выделение избытка углекислого газа организмом, и направленных на поддержание газового гомеостазиса.
Функции и этапы дыхания.
Функции системы дыхания
1.газообмен между клетками организма и окружающей средой
2.выделение летучих соединений
3.депонирование крови
С точки зрения физики газообмен происходит с использованием конвекции (перемещение молекул на большие расстояния с током воздуха и крови) и диффузии (движение газов по градиенту парциального давления на небольшие расстояния).
Этапы (стадии) дыхания
1.Газообмен между внешней средой и альвеолярным воздухом (конвекция)
2.Газообмен между альвеолярным воздухом и кровью (диффузия)
3.Транспорт газов кровью по малому и большому кругу кровообращения (конвекция)
4.Газообмен в тканях (диффузия)
5.Клеточное дыхание (изучает биохимия)
Газообмен в легких происходит благодаря ритмичным дыхательным движениям, инспирации (вдох) и экспирации (выдох). Длительность фазы вдоха и выдоха при различных нагрузках на организм меняется, поэтому введено понятие о паттерне дыхания.
Паттерн дыхания – это совокупность объемных и временных параметров, характеризующих структуру дыхательного цикла и легочную вентиляцию в целом.
Параметры дыхания.
1.Количество дыхательных циклов в 1 минуту. Частота дыхания.
2.Длительность одного дыхательного цикла.
3.Длительность инспираторной и экспираторной фазы.
4.Дыхательный объем или глубина дыхания.
5.Легочная вентиляция (минутный объем дыхания)
Выделяют
нормопноэ , или нормопноическое дыхание, (12-16 дыхательных циклов в мин);
тахипноэ (частое, но неглубокое дыхание, более 20 циклов в минуту);
брадипноэ (медленное, глубокое дыхание, менее 8 вдохов-выдохов в минуту
Рабочее гиперпноэ может наблюдаться при мышечной нагрузке.
Необходимую для организма интенсивность альвеолярной вентиляции можно обеспечить при различных паттернах дыхания, частоты и глубины его.
Чем больше дыхательный объем, тем большее усилие необходимо приложить для преодоления эластичной тяги легких, т.е. при таком дыхании большая нагрузка ложится на вдыхательные мышцы. С другой стороны, при частом поверхностном дыхании нагрузка на дыхательную мускулатуру возрастает из-за сопротивления току воздуха в воздухоносных путях.
При физиологической одышке может быть частое поверхностное дыхание, встречается такой паттерн дыхания при повышенной температуре воздуха и гипертермии. Газообмен в этом случае происходит только в пределах мертвого пространства, отчего обмен кислорода и диоксида углерода в альвеолах снижен.
Кроме указанных, различают понятия гиперпноэ и гипервентиляция легких, в первом случае газообмен в альвеолах нормален, во втором происходит «вымывание» СО 2 из альвеол, и из крови, наступаетгипокапния. При гиповентиляции наблюдаетсягиперкапния , избыток углекислоты в крови или альвеолярном газе.
Недостаток кислорода обозначается как гипоксия , недостаток кровоснабжения в тканях –ишемия .
Дыхательные движения обеспечиваются работой дыхательных мышц.
Исполнительными (эффекторными) образованиями системы дыхания у человека являются инспираторные и экспираторные мышцы. При сокращении инспираторных мышц объем грудной клетки увеличивается за счет поднятия ребер и уплощения диафрагмы. Основные инспираторы – наружные межреберные мышцы и диафрагма. При вдохе межреберные мышцы подтягивают нижележащие ребра вверх, диафрагма опускается книзу. При глубоком вдохе дополнительно в акт включаются грудино-ключично-сосцевидная и трапециевидная мышцы.
Основные экспираторы – внутренние межреберные мышцы, вспомогательные – мышцы живота. Они способствуют опусканию ребер, а также способствуют пассивному смещению диафрагмы при выдохе.
Грудная клетка герметична. С внутренней стороны она выстлана париетальной плеврой. Между тканью легкого (покрытой висцеральной плеврой) и париетальной плеврой имеется плевральная полость, заполненная плевральной жидкостью.
Клетки париетальной плевры фильтруют до 300 мл плевральной жидкости в час. Висцеральная плевра эту жидкость адсорбирует, причем более активно, чем она секретируется. Этим создаются условия для отрицательного (относительно атмосферного) давления в плевральной полости.
Ткань легкого эластична и стремится занять как можно меньший объем. Поэтому растяжение легких происходит за счет распирающего давления атмосферы, атмосферное давление прижимает легкие к париетальной плевре. Нарушение герметичности грудной клетки носит название пневмоторакса .
Таким образом, периодические экскурсии грудной клетки «затягивают» дыхательную порцию воздуха в трахею и далее в легкие, при условии отрицательного давления в плевральной полости.
Воздухопроводящий путь включает носоглотку, трахею, бронхи, 23 поколения которых составляют бронхиальное дерево.
Кондуктивную зону с общим объемом 130-180 мл состаляют первые 16 поколений бронхов, это анатомическое мертвое пространство, названное так потому, что здесь газообмен с кровью не происходит.
Транзиторная зона , 17-19 ветвления бронхов, может содержать альвеолярные ходы.
Респираторная зона бронхиального дерева включает 20-23-е разветвления бронхов. Бронхи образуют альвеолярные бронхиоли и альвеолы.
Функциональной единицей легких являются дольки. Наиболее мелкие бронхиоли входят в дольку и делятся здесь на 12-18 концевых бронхиолей, те образуют альвеолярные бронхиоли и ацинусы, состоящие из альвеол. Число альвеол у человека варьирует от 300 до 700 миллионов. Общая поверхность доходит до 100-130 кв. метров. Альвеолы густо оплетены капиллярами, куда поступает венозная кровь из легочных артерий, а оксигенированная кровь затем отводится из легочного круга кровообращения по легочным венам в левое предсердие.
Особенности нормальной легочной циркуляции крови заключаются в том, что она обладает низким сосудистым сопротивлением и способна аккумулировать весь минутный объем кровотока, создаваемый правым желудочком сердца. Давление в легочной артерии в фазу систолы равно 20-30 мм рт.ст. Объем крови в легких может составлять до 28% от всей циркулирующей в организме. Только за счет емкостных свойств легочные сосуды могут воспринимать весь кровоток, повышающийся при физической нагрузке, без изменений давления.
Альвеолярный эпителий (респираторный) на поверхности покрыт вырабатывающимся в нем специальным веществом фосфолипопротеиновой природы – сурфактонтом . Пленка сурфактанта уменьшает поверхностное натяжение альвеолярной стенки, что препятсвует слипанию альвеол. Сурфактант постоянно вырабатывается разновидностью эпителиальных клеток – гранулярными пневмоноцитами под контролем блуждающих нервов.
Легочные объемы.
В условиях покоя человек дышит так, что используется только часть всего объема легких, поэтому всегда есть резерв для дополнительного вдоха и выдоха. Но даже при самом глубоком дыхании в легких остается определенное количество воздуха, составляющее остаточный объем .
Общая емкость легких =резервный объем вдоха (2,5 л)+дыхательный объем (500-700 мл)+ резервный объем выдоха (1,5 л) + остаточный объем (1,5 л) =3,5…6 л.
Дыхательный объем – объем воздуха, который входит в легкие при каждом спокойном вдохе и выходит при спокойном выдохе.
Резервные объемы вдоха и выдоха – объемы воздуха, которые человек может произвольно вдохнуть и выдохнуть сверх дыхательного объема.
Жизненная емкость легких – количество воздуха, которое может выдохнуть человек после глубокого вдоха. Она равна сумме дыхательного объема, резервных объемов вдоха и выдоха.
Легочная вентиляция всегда находится в точном соответствии с текущими метаболическими потребностями организма. Увеличение вентиляции происходит как за счет роста дыхательного объема, так и увеличением частоты дыхания.
Не весь воздух, поступающий в легкие, участвует в газообмене, анатомическое мертвое пространство соответствует (в мл) цифре удвоенной массы тела. Функциональное мертвое пространство дополнительно снижает степень газообмена.
Газ в альвеолах имеет постоянный состав, обусловленный буферными функциями мертвого пространства, где воздух увлажняется и нагревается.
В условиях покоя оптимальным является дыхание через нос, хотя при этом сопротивление дыханию возрастает по сравнению с дыханием через рот.
При осуществлении дыхательных движений дыхательные мышцы совершают работу, затрачиваемую на преодоление внутренних и внешних сил. Работа дыхания складывается из энергозатрат на преодоление общего легочного сопротивления (эластичного сопротивления самой легочной ткани и грудной клетки) и преодоления сопротивления потоку воздуха в воздухоносных путях.
Минутному объему дыхания должен соответствовать минутный объем крови, протекающий по сосудам малого круга кровообращения. Вентиляционно-перфузионный коэффициент составляет 0,8-0,9, т. е. при альвеолярной вентиляции, равной 6 л/мин, минутный объем кровообращения может быть равным 7 л/мин.
В атмосфере Земли кислород составляет примерно 21%, или 1/5. Атмосферное давление на уровне моря 760 мм рт.ст. Значит, парциальное давление кислорода примерно соответствует 1/5 этой величины, 160 мм рт.ст., это предельная цифра содержания О 2 в естественных газовых смесях.
В воздухоносных путях воздух постепенно теряет скорость перемещения (конвекции). В респираторных бронхиолях и альвеолах большое значение приобретает диффузия газов. Газы перемещаются по градиенту парциального давления. В альвеолах, где и происходит, контакт альвеолярного газа с капиллярной кровью, напряжение кислорода Р О 2 составляет 103 мм рт.ст., а парциальное давление диоксида углерода Р СО 2 около 40 мм рт.ст. В выдыхаемом воздухе Р О 2 составляет 126 мм рт.ст., а Р СО 2 соответственно 16 мм рт.ст. В артериальной крови Р О 2 соответствует 95 мм рт.ст., в венозной Р О 2 равно 40 мм рт.ст. Р СО 2 артериальной крови соответствует 40 мм рт.ст., а венозной – Р СО 2 приближается к 46 мм рт.ст.
Вектор диффузии дыхательных газов
Поэтому вектор диффузии кислорода постоянно направлен в сторону альвеол и капилляров, а углекислоты – в обратном направлении, из капилляров в атмосферу.
Перенос кислорода из альвеолярного газа в кровь и диоксида углерода из крови в альвеолярный газ происходит исключительно путем диффузии. Движущей силой диффузии диффузии служит градиент парциального давления каждого из газов по обе стороны аэрогематического барьера. Диффузия осуществляется в водной среде. В слое сурфактанта растворимость кислорода повышается.
Аэрогематический барьер состоит из слоя сурфактанта, альвеолярного эпителия, двух основных мембран, эндотелия капилляра и мембраны эритроцита.
Диффузионная способность легких для кислорода достаточно высока. Установлено, что на каждый миллиметр ртутного столба градиента парциального давления кислорода между альвеолярным газом и эритроцитом в кровь поступает путем диффузии 25 мл кислорода в минуту. Этого достаточно для того, чтобы за 0,8 с, что равно времени прохождения отдельным эритроцитом одного легочного капилляра, парциальное давление кислорода в нем успело выравняться с альвеолярным. Даже с большим запасом по времени, поскольку для выравнивания напряжения кислорода в эритроцитах с альвеолярным воздухом достаточно 0,25 с.
Поэтому, если кровоток в капиллярах легких повышается (возрастает линейная скорость движения эритроцитов) при физической нагрузке на организм, и время прохождения капилляров клетками уменьшается до 0,3 с, этого оказывается вполне достаточно для полного газообмена. Для диффузии из крови углекислого газа необходимо всего 0,1 с. Растворимость диоксида углерода в воде превышает этот показатель для кислорода в 25 раз.
Транспорт кислорода кровью.
Если животное имеет систему кровообращения, в крови имеется переносчик кислорода. В растворенном состоянии у человека в артериальной крови имеется только 2% кислорода.
Все пигменты – переносчики кислорода представляют собой металлорганические соединения, большинство содержит Fe, некоторые Cu.
Гемоглобины представляют собой железопорфирины (гем), связанные с глобином (белком). Гемоглобин у человека и млекопитающих всегда находится в специализированных клетках крови эритроцитах. Установлено более 90 типов гемоглобинов, отличающихся белковыми составляющими. Молекула гемоглобина состоит из нескольких мономеров, каждый из которых содержит один гем, соединенный с глобином. У человека гемоглобин содержит 4 таких мономера. Миоглобин содержит только 1 гем.
Гем в химическом отношении представляет собой протопорфирин, состоящий из 4 пиррольных колец с атомом железа в центре.
Оксигенация гемоглобина представляет собой обратимое присоединение кислорода к закисному (двухвалентному) железу в количествах, зависящих от напряжения кислорода в окружающем пространстве.
Кислород присоединяется к каждому из атомов железа согласно уравнению равновесия
Формально в этой реакции не происходит изменения валентности железа. Тем не менее оксигенация сопровождается частичным переходом электрона от закисного железа к кислороду, кислород частично восстанавливается.
Иное значение валентности может быть у гемового железа при образовании метгемоглобина, когда Fe изменяет валентность и становится трехвалентным. В этом случае, при истинном окислении железа, гемоглобин утрачивает способность переносить кислород.
Гем в молекуле гемоглобина способен присоединять другие молекулы. Если он присоединяет диоксид углерода, его называют карбогемоглобином. Если к гему присоединятся монооксид углерода, образуется карбоксигемоглобин. Сродство гемоглобина к CO в 300 раз выше, чем к О 2 . Поэтому отравление угарным газом очень опасно. Если во вдыхаемом воздухе содержится 1% СО, млекопитающие и птицы могут погибнуть.
Артериальная кровь насыщается кислородом на 96-97%. Этот процесс происходит очень быстро, всего за четверть секунды в альвеолярных капиллярах.
В литературе принято оценивать содержание кислорода в крови по показателю кислородная емкость крови .
Кислородная емкость крови – это максимальное количество кислорода, которое может присоединить 100 мл крови.
Поскольку 96% кислорода находится в соединении с гемоглобином, кислородная емкость крови определяется этим пигментом. Известно, что кислород-связывающая способность 1 г гемоглобина определяется величиной 1,34 – 1,36 мл О 2 , при нормальном атмосферном давлении. Это означает, что при содержании в крови 15 г% Нв (а это близко к средней), кислородная емкость составляет 1,341520 объемных процентов, то есть на каждые 20 мл О 2 на каждые 100 мл крови, или 200 мл О 2 на литр крови. В 5 литрах крови (полная кислородная емкость индивидуума, у которого 5 л крови в системе кровообращения) содержится 1 литр кислорода.
Реакция оксигенации гемоглобина обратима
HHb 4 +4O 2 = HHb 4 (O 2) 4
Или проще Hb+О 2 = HbО 2
Оказалось, что на практике удобнее анализировать этот процесс, если построить график зависимости концентрации HbО 2 в образце от парциального давления/напряжения кислорода. Чем больше в среде кислорода, тем сильнее равновесие в реакции смещается в сторону оксигенации, и наоборот.
Каждому значению РО 2 соответствует определенный процент HbО 2 . При значениях РО 2 , характерных для артериальной крови, практически весь гемоглобин окислен. В периферических тканях, при низких значениях напряжения кислорода, увеличивается скорость диссоциации его диссоциации до кислорода и гемоглобина.
Кривая диссоциации гемоглобина имеется в каждом учебнике.
Анализ кривой диссоциации оксигемоглобина показывает, что при напряжении кислорода в среде 60-100 мм рт.ст. (условия равнины и подъема человека на высоту до 2 километров) насыщение кислородов крови происходит полностью. В тканях отдача кислорода также протекает удовлетворительно, при напряжениях кислорода около 20 мм рт.ст.
Другими словами, характер кривой дает сведения о свойствах транспортной системы.
Диссоциация оксигемоглобина зависит не только от парциального давления кислорода в тканях, но и от некоторых других условий. Когда в кровь поступает углекислота из тканей, сродство гемоглобина к кислороду падает и кривая диссоциации сдвигается вправо. Это прямой эффект Вериго-Бора. Эффект Вериго-Бора способствует улучшению диссоциации оксигемоглобина в тканях. Обратный эффект наблюдается в легких, где отдача диоксида углерода приводит к более полному насыщению гемоглобина кислородом. Эффект обусловлен не самим СО 2 , а подкислением среды при образовании угольной кислоты (или накоплением молочной кислоты в активно работающих мышцах).
Не весь оксигемоглобин диссоциирует в тканях. От 40 до 70% его сохраняется в венозной крови. У человека каждые 100 мл крови отдают тканям 5-6 мл кислорода, и на такую же величину обогащаются новой его порцией в легочных капиллярах. Для оценки этих процессов (утилизации кислорода тканями) введен показатель артерио-венозная разница по кислороду.
Транспорт углекислого газа кровью.
Как и кислород, диоксид углерода в крови находится в двух состояниях – физически растворенном и химически связанном. Около 5% СО 2 транспортируется в растворенном виде. Химическая связь диоксида углерода осуществляется по реакции
СО 2 + Н 2 О↔Н 2 СО 3 ↔Н + + НСО 3 -
Реакция сдвигается вправо при высоких напряжениях СО 2 , и влево при низких. Катализируется карбоангидразой с коэффициентом ускорения 250-300 раз. 80% образовавшейся угольной кислоты транспортируется в виде бикарбонатов щелочных металлов. Противоионами для карбонатных ионов в плазме выступают Na + , а в эритроитах – К + . Остальные 20% НСО 3 - транспортируется в связи с гемоглобином. В артериальной крови 15% СО 2 (в венозной 20%) переносится в виде карбаминовых групп гемоглобина, поскольку NH-группы белка связывают СО 2 обратимо. Доля транспорта в связи с гемом ничтожна. Для образования бикарбонатов щелочных металлов используется их резерв, ассоциированный с молекулой гемоглбина. Гемоглобин, как и все белковые молекулы, является амфотерным соединением. В слабощелочной среде (рН 7,35-7,4) гемоглобин и оксигемоглобин ведут себя как слабые кислоты, ассоциируя ионы калия. В артериальной крови 67% НСО 3 - растворено в плазме, уравновешивающим ионом выступает натрий. Гемоглобин в дезоксиформе слабее по кислотным свойствам, чем оксигемоглобин, поэтому легко отдает К + , при этом реализуется эффект Холдена: оксигенация гемоглобина облегчает отдачу СО 2 кровью, а дезоксигенация гемоглобина усиливает поглощение диоксида углерода.
Гемоглобиновая буферная система (отвечает за 75% буферных свойств крови) и карбонатная буферная система, кроме дыхательной функции, обеспечивает постоянство активной реакции крови в диапазоне рН 7,35 – 7,47 (венозная кровь в норме имеет более кислую реакцию на 0,02 единицы рН). Поэтому нарушения кислотно-щелочного равновесия в организме может быть не только метаболическим, но и дыхательным. При респираторном ацидозе рН крови снижается, концентрация НСО 3 - возрастает. При респираторном алкалозе (может наступить при гипервентиляции) наблюдаются обратные процессы, концентрация НСО 3 - падает из-за «вымывания» углекислого газа.
Регуляция дыхания.
Конечная цель регуляции дыхания, или полезный приспособительный результат – поддержание постоянного газового состава и рН артериальной и венозной крови. Отклонение этих показателей от нормы (РО 2 менее 100 мс рт.ст., РСО 2 более 40 мм рт.ст., рН от 7,36) воспринимается как стимул для регуляции. Координированные сокращения дыхательных мышц обеспечиваются ритмической активностью нейронов дыхательного центра, или, как это принято формулировать по современным представлениям, центрального дыхательного механизма .
К дыхательным нейронам относят те нервные клетки, импульсная активность которых меняется в соответствии с фазами дыхательного цикла. Различают инспираторные нейроны (нейроны вдоха) и экспираторные (нейроны выдоха) и клеточные популяции, согласовывающие смену дыхательных фаз. Центральный дыхательный механизм локализован в ретикулярной формации продолговатого мозга. Большинство нейронов сгруппированы в двух главных группах ядер – дорсальной и вентральной. В дорсальной группе сосредоточены инспираторные нейроны, посылающие аксоны в шейные сегменты спинного мозга, где они синаптически оканчиваются на мотонейронах ядра диафрагмального нерва. Ядра вентральной группы дыхательных ядер содержат как инспираторные, так и экспираторные нейроны. Они связаны синаптически с теми нейронами спинного мозга, которые иннервируют межреберные мышцы. Для 80% нейронов грудного отдела спинного мозга характерна дыхательная ритмика. В области моста выделен пневмотаксический центр, клетки которого принимают участие в переключении фаз дыхательного цикла. Для нейронов центрального дыхательного механизма характерен автоматизм, хотя пейсмекеров пока не обнаружено. Основной активатор дыхательного механизма – афферентная сигнализация от рецепторов, расположенных во внутренней среде организма. Главный дыхательный стимул – снижение в крови содержания кислорода и повышение напряжения диоксида углерода. Хеморецепторы посылают в ЦНС сигналы о степени отклонения этих показателей от нормы. Основное место локализации хеморецепторов дыхательной системы – область каротидного синуса (каротидные клубочки). В области дуги аорты расположена вторая группа хеморецепторов, контролирующая газовые и кислотные показатели той порции крови, которая направляется к внутренним органам. В продолговатом мозге имеются и центральные хеморецепторы.
Установлено, что чем выше в крови рСО 2 , тем выше частота импульсации в афферентных волокнах синокаротидного нерва. Эта афферентная посылка интегрируется центральным дыхательным механизмом и используется для усиления дыхания, как увеличением частоты дыхательных циклов, так и углублением каждого вдоха.
В трахее, бронхах имеются собственные рецепторы, инициирующие защитные рефлексы дыхания, например, кашель. Кроме того, часть из них используется и для коррекции частоты и глубины дыхания. К ним относится рефлекс Геринга–Брейера. Рецепторы, реагирующие на повышение давления в воздухоносных путях, активируются при вдохе и посылают афферентные сигналы по волокнам блуждающего нерва к группе нейронов дорсальной порции дыхательного центра. Их возбуждение нарастает в фазу вдоха и тормозит инспираторные нейроны. Каждый вдох за счет рецепторов растяжения подготавливает свое окончание.
Имеются рецепторы и в верхних дыхательных путях, они активируются при попадании в нос и рот пыли или ирритантов. Кашель, чихание, принюхивание, остановка дыхания на вдохе при обнаружении резкого неприятного запаха или химическом загрязнении среды – рефлекторные проявления их активации.
человека (газообмен между вдыхаемым атмосферным воздухом и циркулирующей по малому кругу кровообращения кровью ).Введение
Дыхание является одной из важнейших функций регулирования жизнедеятельности человеческого организма.
В организме человека функцию дыхания обеспечивает дыхательная (респираторная система).
В дыхательную систему входят легкие и респираторный тракт (дыхательные пути), который, в свою очередь, включает носовые ходы, гортань, трахею, бронхи, мелкие бронхи и альвеолы. Бронхи разветвляются, распространяясь по всему объему легких, и напоминают крону дерева. Поэтому часто трахею и бронхи со всеми ответвлениями называют бронхиальным деревом .
Основная функция дыхательной системы заключается в обеспечении газообмена О2 и СО2 между окружающей средой и организмом в соответствии с его метаболическими потребностями. В целом эту функцию регулирует сеть многочисленных нейронов центральной нервной системы (ЦНС), которые связаны с дыхательным центром продолговатого мозга .
Газообмен осуществляется в альвеолах лёгких , и в норме направлен на захват из вдыхаемого воздуха кислорода и выделение во внешнюю среду образованного в организме углекислого газа .
Взрослый человек, находясь в состоянии покоя, совершает в среднем 14 дыхательных движений в минуту, однако частота дыхания может претерпевать значительные колебания (от 10 до 18 за минуту) . Взрослый человек делает 15-17 вдохов-выдохов в минуту, а новорождённый ребёнок делает 1 вдох в секунду. Вентиляция альвеол осуществляется чередованием вдоха (инспирация) и выдоха (экспирация). При вдохе в альвеолы поступает атмосферный воздух , а при выдохе из альвеол удаляется воздух, насыщенный углекислым газом.
Общая характеристика дыхания
По способу расширения грудной клетки различают два типа дыхания:
- грудной тип дыхания (расширение грудной клетки производится путём поднятия рёбер), чаще наблюдается у женщин;
- брюшной тип дыхания (расширение грудной клетки производится путём уплощения диафрагмы ), чаще наблюдается у мужчин .
По функционированию различают:
- внешнее дыхание - это поступление кислорода в легкие и газообмен между воздухом альвеол и кровью малого круга;
- внутреннее дыхание - утилизация кислорода в тканях, т. е. его участие в окислительно-восстановительных реакциях. Этот процесс протекает в митохондриях. Внутренне дыхание изучается в курсе биохимии.
Между внешним и внутренним дыханием имеется промежуточное звено - транспорт газов кровью. Обеспечивается не дыхательной системой, а сердечно-сосудистой системой и системой крови .
Дыхание-совокупность последовательно протекающих процессов, обеспечивающих потребление организмом О2 и выделение СО2.
Кислород поступает в составе атмосферного воздуха в легкие, транспортируется кровью и тканевыми жидкостями к клеткам и используется для биологического окисления. В процессе окисления образуется двуокись углерода, которая поступает в жидкие среды организма, транспортируется ими в легкие и выводится в окружающую среду.
Дыхание включает следующие процессы (этапы):
- обмен воздуха между внешней средой и альвеолами легких (внешнее дыхание, или вентиляция легких);
- обмен газов между альвеолярным воздухом и кровью, протекающей через легочные капилляры (диффузия газов в легких);
- транспорт газов кровью;
- обмен газов между кровью и тканями в тканевых капиллярах (диффузия газов в тканях);
- потребление кислорода клетками и выделение ими углекислого газа (клеточное дыхание).
На рисунке 1 представлена схема легочного пузырька и газообмена в легких.
Рисунок 1 – Легочный пузырек. Газообмен в легких .
В дыхательных путях газообмен не происходит, и состав воздуха не меняется. Пространство, заключенное в дыхательных путях называется мертвым, или вредным. При спокойном дыхании объем воздуха в мертвом пространстве составляет 140-150 мл.
Предметом рассмотрения физиологии являются первые 5 процессов. Внешнее дыхание осуществляется благодаря изменениям объема грудной полости, влияющим на объем легких.
Объем грудной полости увеличивается во время вдоха (инспирация) и уменьшается во время выдоха (экспирация). Легкие пассивно следуют за изменениями объема грудной полости, расширяясь при вдохе и спадаясь при выдохе. Эти дыхательные движения обеспечивают вентиляцию легких за счет того, что при вдохе воздух по воздухоносным путям поступает в альвеолы, а при выдохе покидает их. Изменение объема грудной полости осуществляется в результате сокращений дыхательных мышц.
Дыхательный цикл состоит из двух фаз - вдох и выдох. Соотношение вдоха и выдоха - 1: 1,2.
Важнейший механизм газообмена – это диффузия , при которой молекулы перемещаются из области их высокого скопления в область низкого содержания без затраты энергии (пассивный транспорт ). Перенос кислорода из окружающей среды к клеткам производится путем транспорта кислорода в альвеолы, далее в кровь. Таким образом, венозная кровь обогащается кислородом и превращается в артериальную. Поэтому состав выдыхаемого воздуха отличается от состава наружного воздуха: в нем содержится меньше кислорода и больше углекислого газа, чем в наружном, и много водяных паров. Кислород связывается с гемоглобином , который содержится в эритроцитах, насыщенная кислородом кровь поступает в сердце и выталкивается в большой круг кровообращения. По нему кровь разносит кислород по всем тканям организма. Поступление кислорода в ткани обеспечивает их оптимальное функционирование, при недостаточном же поступлении наблюдается процесс кислородного голодания (гипоксии ).
Недостаточное поступление кислорода может быть обусловлено несколькими причинами как внешними (уменьшение содержания кислорода во вдыхаемом воздухе), так и внутренними (состояние организма в данный момент времени). Пониженное содержание кислорода во вдыхаемом воздухе, так же как и увеличение содержания углекислого газа и других вредных токсических веществ наблюдается в связи с ухудшением экологической обстановки и загрязнением атмосферного воздуха. По данным экологов только 15% горожан проживают на территории с допустимым уровнем загрязнения воздуха, в большинстве же районов содержание углекислого газа увеличено в несколько раз.
При очень многих физиологических состояниях организма (подъем в гору, интенсивная мышечная нагрузка), так же как и при различных патологических процессах (заболевания сердечно-сосудистой, дыхательной и других систем) в организме также может наблюдаться гипоксия.
Природа выработала множество способов, с помощью которых организм приспосабливается к различным условиям существования, в том числе к гипоксии. Так компенсаторной реакцией организма, направленной на дополнительное поступление кислорода и скорейшее выведение избыточного количества углекислого газа из организма является углубление и учащение дыхания. Чем глубже дыхание, тем лучше вентилируются легкие и тем больше кислорода поступает к клеткам тканей.
К примеру, во время мышечной работы усиление вентиляции легких обеспечивает возрастающие потребности организма в кислороде. Если в покое глубина дыхания (объем воздуха, вдыхаемого или выдыхаемого за один вдох или выдох) составляет 0,5 л, то во время напряженной мышечной работы она увеличивается до 2-4 л в 1 минуту. Расширяются кровеносные сосуды легких и дыхательных путей (а также дыхательных мышц), увеличивается скорость тока крови по сосудам внутренних органов. Активируется работа дыхательных нейронов. Кроме того, в мышечной ткани есть особый белок (миоглобин ), способный обратимо связывать кислород. 1 г миоглобина может связать примерно до 1,34 мл кислорода. Запасы кислорода в сердце составляют около 0,005 мл кислорода на 1 г ткани и этого количества в условиях полного прекращения доставки кислорода к миокарду может хватить для того, чтобы поддерживать окислительные процессы лишь в течение примерно 3-4 с.
Миоглобин играет роль кратковременного депо кислорода. В миокарде кислород, связанный с миоглобином, обеспечивает окислительные процессы в тех участках, кровоснабжение которых на короткий срок нарушается.
В начальном периоде интенсивной мышечной нагрузки увеличенные потребности скелетных мышц в кислороде частично удовлетворяются за счет кислорода, высвобождающегося миоглобином. В дальнейшем возрастает мышечный кровоток, и поступление кислорода к мышцам вновь становится адекватным.
Все эти факторы, включая усиление вентиляции легких, компенсируют кислородный “долг”, который наблюдается при физической работе. Естественно, увеличению доставки кислорода к работающим мышцам и удалению углекислого газа способствует согласованное увеличение кровообращения в других системах организма.
Мышечное обеспечение дыхания
Дыхательные мышцы обеспечивают ритмичное увеличение или уменьшение объема грудной полости. Функционально дыхательные мышцы делят на инспираторные (основные и вспомогательные) и экспираторные.
Основную инспираторную группу мышц составляют диафрагма, наружные межреберные и внутренние межхрящевые мышцы; вспомогательные мышцы - лестничные, грудиноключично-сосцевидные, трапецевидная, большая и малая грудные мышцы. Экспираторную группу мышц составляют абдоминальные (внутренняя и наружная косые, прямая и поперечная мышцы живота) и внутренние межреберные.
Важнейшей мышцей вдоха является диафрагма - куполообразная поперечнополосатая мышца, разделяющая грудную и брюшную полости. Она прикрепляется к трем первым поясничным позвонкам (позвоночная часть диафрагмы) и к нижним ребрам (реберная часть). К диафрагме подходят нервы от III-V шейных сегментов спинного мозга. При сокращении диафрагмы органы брюшной полости смещаются вниз и вперед и вертикальные размеры грудной полости возрастают.
Кроме того, при этом поднимаются и расходятся ребра, что приводит к увеличению поперечного размера грудной полости. При спокойном дыхании диафрагма является единственной активной инспираторной мышцей и ее купол опускается на 1 -1,5 см.
Известно два биомеханизма, которые изменяют объем грудной клетки: поднятие и опускание ребер и движения купола диафрагмы; оба биомеханизма осуществляются дыхательными мышцами. Дыхательные мышцы подразделяют наинспираторные и экспираторные.
Экспираторными мышцами являются внутренние межреберные и мышцы брюшной стенки, или мышцы живота. Последние нередко относят к главным экспираторным мышцам. У нетренированного человека они участвуют в дыхании при вентиляции легких свыше 40 л*мин-1.
Движения ребер. Каждое ребро способно вращаться вокруг оси, проходящей через две точки подвижного соединения с телом я поперечным отростком соответствующего позвонка.
Сокращение этих мышц вызывает перемещение ребер, что оказывает
содействие инспираторным мышцам. При спокойном дыхании вдох осуществляется активно, а выдох пассивно. Силы, обеспечивающие спокойный выдох:
- сила тяжести грудной клетки
- эластическая тяга легких
- давление органов брюшной полости
- эластическая тяга перекрученных во время вдоха реберных хрящей
В активном выдохе принимают участие внутренние межреберные мышцы, задняя нижняя зубчатая мышца, мышцы живота
осуществление форсированного дыхания;
При глубоком форсированном дыхании увеличивается амплитуда движений диафрагмы (экскурсия может достигать 10 см) и активизируются наружные межреберные и вспомогательные мышцы. Из вспомогательных мышц наиболее значимыми являются лестничные и грудиноключично-сосцевидные мышцы.
Наружные межреберные мышцы соединяют соседние ребра. Их волокна ориентированы наклонно вниз и вперед отверхнего к нижнему ребру. При сокращении этих мышц ребра поднимаются и смещаются вперед, что приводит к увеличению объема грудной полости в переднезаднем и боковом направлениях. Паралич межреберных мышц не вызывает серьезных расстройств дыхания, поскольку диафрагма обеспечивает вентиляцию.
Лестничные мышцы, сокращаясь во время вдоха, поднимают 2 верхних ребра, а вместе сними всю грудную клетку. Грудинно-ключично-сосцевидные мышцы поднимают I ребро и грудину. При спокойном дыхании они практически не задействованы, однако при увеличении легочной вентиляции могут интенсивно работать.
Величина давления в плевральной полости и легких при дыхании
Давление в герметично замкнутой плевральной полости между висцеральным и париетальным листками плевры зависит от величин и направления сил, создаваемых эластической паренхимой легких и грудной стенкой. Плевральное давление можно измерить манометром, соединенным с плевральной полостью полой иглой. В клинической практике часто применяют косвенный метод оценки величины плеврального давления, измеряя давление в нижней части пищевода с помощью пищеводного баллонного катетера. Внутри пищеводное давление во время дыхания отражает изменения внутри плеврального давления.
Плевральное давление ниже атмосферного во время вдоха, а во время выдоха может быть ниже, выше или равным атмосферному в зависимости от форсированности выдоха. При спокойном дыхании плевральное давление перед началом вдоха составляет -5 см вод.ст., перед началом выдоха оно понижается еще на 3-4 см вод.ст. При пневмотораксе (нарушение герметичности грудной клетки и сообщение плевральной полости с внешней средой) выравниваются плевральное и атмосферное давления, что вызывает спадение легкого и делает невозможной его вентиляцию.
Значение сурфактанта:
- создает возможность расправления легкого при первом вдохе новорожденного;
- препятствует развитию ателектаза при выдохе;
- обеспечивает до ⅔ эластического сопротивления ткани легкого взрослого человека и стабильность структуры респираторной зоны;
- регулирует скорость адсорбции О2 по границе раздела фаз газ – жидкость и интенсивность испарения Н2О с альвеолярной поверхности;
- очищает поверхность альвеол от попавших с дыханием инородных частиц и обладает бактериостатической активностью.
Саморегуляция дыхания.
Организм осуществляет тонкое регулирование содержания кислорода и углекислого газа в крови, которое остается относительно постоянным, несмотря на колебания количества поступающего кислорода и потребности в нем. Во всех случаях регуляция интенсивности дыхания направлена на конечный приспособительный результат – оптимизацию газового состава внутренней среды организма.
Частота и глубина дыхания регулируются нервной системой – ее центральными (дыхательный центр ) и периферическими (вегетативными) звеньями. В дыхательном центре, расположенном в головном мозге, имеются центр вдоха и центр выдоха.
Дыхательный центр представляет совокупность нейронов, расположенных в продолговатом мозге центральной нервной системы.
При нормальном дыхании центр вдоха посылает ритмические сигналы к мышцам груди и диафрагме, стимулируя их сокращение. Ритмические сигналы образуются в результате спонтанного образования электрических импульсов нейронами дыхательного центра.
Сокращение дыхательных мышц приводит к увеличению объема грудной полости, в результате чего воздух входит в легкие. По мере увеличения объема легких возбуждаются рецепторы растяжения, расположенные в стенках легких; они посылают сигналы в мозг – в центр выдоха. Этот центр подавляет активность центра вдоха, и поток импульсных сигналов к дыхательным мышцам прекращается. Мышцы расслабляются, объем грудной полости уменьшается, и воздух из легких вытесняется наружу (рисунок 2).

Рисунок 2- Регуляция дыхания
Процесс дыхания, как уже отмечалось, состоит из легочного (внешнего) дыхания, а также транспорта газа кровью и тканевого(внутреннего) дыхания. Если клетки организма начинают интенсивно использовать кислород и выделять много углекислого газа, то в крови повышается концентрация угольной кислоты. Кроме того, увеличивается содержание молочной кислоты в крови за счет усиленного образования ее в мышцах. Данные кислоты стимулируют дыхательный центр, и частота и глубина дыхания увеличиваются. Это еще один уровень регуляции. В стенках крупных сосудов, отходящих от сердца, имеются специальные рецепторы, реагирующие на понижение уровня кислорода в крови. Эти рецепторы также стимулируют дыхательный центр, повышая интенсивность дыхания. Данный принцип автоматической регуляции дыхания лежит в основе бессознательного управления дыханием, что позволяет сохранить правильную работу всех органов и систем независимо от условий, в которых находится организм человека.
Ритмичность дыхательного процесса, различные типы дыхания. В норме дыхание представлено равномерными дыхательными циклами “вдох – выдох” до 12-16 дыхательных движений в минуту. В среднем такой акт дыхания совершается за 4-6 с. Акт вдоха проходит несколько быстрее, чем акт выдоха (соотношение длительности вдоха и выдоха в норме составляет 1:1,1 или 1:1,4). Такой тип дыхания называется эйпноэ (дословно – хорошее дыхание). При разговоре, приеме пищи ритм дыхания временно меняется: периодически могут наступать задержки дыхания на вдохе или на выходе (апноэ ). Во время сна также возможно изменение ритма дыхания: в период медленного сна дыхание становится поверхностным и редким, а в период быстрого – углубляется и учащается. При физической нагрузке за счет повышенной потребности в кислороде возрастает частота и глубина дыхания, и, в зависимости от интенсивности работы, частота дыхательных движений может достигать 40 в минуту.
При смехе, вздохе, кашле, разговоре, пении происходят определенные изменения ритма дыхания по сравнению с так называемым нормальным автоматическим дыханием. Из этого следует, что способ и ритм дыхания можно целенаправленно регулировать с помощью сознательного изменения ритма дыхания.
Человек имеет возможность сознательно управлять дыханием.
Человек рождается уже с умением использовать лучший способ дыхания. Если проследить как дышит ребенок, становится заметным, что его передняя брюшная стенка постоянно поднимается и опускается, а грудная клетка остается практически неподвижной. Он “дышит” животом – это так называемый диафрагмальный тип дыхания.
Диафрагма – это мышца, разделяющая грудную и брюшную полости. Сокращения данной мышцы способствуют осуществлению дыхательных движений: вдоха и выдоха.
В повседневной жизни человек не задумывается о дыхании и вспоминает о нем, когда по каким-то причинам становится трудно дышать. Например, в течение жизни напряжение мышц спины, верхнего плечевого пояса, неправильная осанка приводят к тому, что человек начинает “дышать” преимущественно только верхними отделами грудной клетки, при этом объем легких задействуется всего лишь на 20%. Попробуйте положить руку на живот и сделать вдох. Заметили, что рука на животе практически не изменила своего положения, а грудная клетка поднялась. При таком типе дыхания человек задействует преимущественно мышцы грудной клетки (грудной тип дыхания) или области ключиц (ключичное дыхание). Однако как при грудном, так и при ключичном дыхании организм снабжается кислородом в недостаточной степени.
Недостаток поступления кислорода может возникнуть также при изменении ритмичности дыхательных движений, то есть изменении процессов смены вдоха и выдоха.
В состоянии покоя кислород относительно интенсивно поглощается миокардом, серым веществом головного мозга (в частности, корой головного мозга), клетками печени и корковым веществом почек; клетки скелетной мускулатуры, селезенка и белое вещество головного мозга потребляют в состоянии покоя меньший объем кислорода, то при физической нагрузке потребление кислорода миокардом увеличивается в 3-4 раза, а работающими скелетными мышцами – более чем в 20-50 раз по сравнению с покоем.
Интенсивное дыхание, состоящее в увеличении скорости дыхания или его глубины (процесс называется гипервентиляцией ), приводит к увеличению поступления кислорода через воздухоносные пути. Однако частая гипервентиляция способна обеднить ткани организма кислородом. Частое и глубокое дыхание приводит к уменьшению количества углекислоты в крови (гипокапнии ) и защелачиванию крови – респираторному алкалозу.
Подобный эффект прослеживается, если нетренированный человек осуществляет частые и глубокие дыхательные движения в течение короткого времени. Наблюдаются изменения со стороны как центральной нервной системы (возможно появление головокружения, зевоты, мелькания “мушек” перед глазами и даже потери сознания), так и сердечно-сосудистой системы (появляется одышка, боль в сердце и другие признаки). В основе данных клинических проявлений гипервентиляционного синдрома лежат гипокапнические нарушения, приводящие к уменьшению кровоснабжения головного мозга. В норме у спортсменов в покое после гипервентиляции наступает состояние сна.
Следует отметить, что эффекты, возникающие при гипервентиляции, остаются в то же время физиологичными для организма – ведь на любое физическое и психоэмоциональное напряжение организм человека в первую очередь реагирует изменением характера дыхания.
При глубоком, медленном дыхании (брадипноэ ) наблюдается гиповентиляционный эффект. Гиповентиляция – поверхностное и замедленное дыхание, в результате которого в крови отмечается понижение содержание кислорода и резкое увеличение содержания углекислого газа (гиперкапния ).
Количество кислорода, которое клетки используют для окислительных процессов, зависит от насыщенности крови кислородом и степени проникновения кислорода из капилляров в ткани.Снижение поступления кислорода приводит к кислородному голоданию и к замедлению окислительных процессов в тканях.
В 1931 году доктор Отто Варбург получил Нобелевскую премию в области медицины, открыв одну из возможных причин возникновения рака. Он установил, что возможной причиной этого заболевания является недостаточный доступ кислорода к клетке.
Правильное дыхание, при котором воздух, проходящий через воздухоносные пути, в достаточной степени согревается, увлажняется и очищается – это спокойное, ровное, ритмичное, достаточной глубины.
Во время ходьбы или выполнения физических упражнений следует не только сохранять ритмичность дыхания, но и правильно сочетать ее с ритмом движения (вдох на 2-3 шага, выдох на 3-4 шага).
Важно помнить, что потеря ритмичности дыхания приводит к нарушению газообмена в легких, утомлению и развитию других клинических признаков недостатка кислорода.
При нарушении акта дыхания уменьшается приток крови к тканям и понижается насыщение ее кислородом.
Необходимо помнить, что физические упражнения способствуют укреплению дыхательной мускулатуры и усиливают вентиляцию легких. Таким образом, от правильного дыхания в значительной мере зависит здоровье человека .
Физиология дыхательных путей
Регуляция величины просвета бронхов.
Гладкие мышци бронхиол иннервируются волокнами вегетативной нервной системы. Прямое влияние симпатической системы незначительное, зато катехоламины, которые находятся в крови, особенно адреналин, действуя на b-адренорецепторы, оказывает расслабление этих мышц.
Ацетилхолин, который выделяется волокнами блуждающего нерва, суживает бронхиолы. Поэтому введение атропина сульфата может вызвать расширение бронхиол. При участие парасимпатических нервов реализируется ряд рефлексов, которые начинаются в дыхательных путях в случае раздражения их рецепторов дымом, отравляющими газами, инфекцией т.п.. Некоторые вещества, которые осуществляют аллергические реакции, также могут суживать бронхиолы .
Список литературы
- Научно популярное методическое пособие «Дыхательная система. Физиология дыхания» [Электронный ресурс ].- Режим доступа: http://www.rlsnet.ru/books_book_id_2_page_30.htm
- Свободная электронная энциклопедия
- Обсуждение «Внутреннее и внешнее дыхание. Их отличие» [Электронный ресурс ].- Режим доступа: http://otvet.mail.ru/question/49261280
Российский Государственный Университет Физической Культуры, Спорта, Молодёжи и Туризма
Факультет "Физическая культура и спорт"
Отделения "ТиМ художественной гимнастики"
Реферат по дисциплине "Физиология"
на тему: "Физиология дыхания"
Выполнила:
Студентка 3 курса
Малахова Е.В.
Преподаватель:
Захарьева Н.Н.
Москва 2011-
Введение
1. Характеристика понятия "дыхание"
2. Внешнее дыхание
3. Легочная вентиляция
4. Обмен газов в легких и их перенос
5. Регуляция дыхания
6. Особенности дыхания при физической нагрузке и при измененном парциальном давлении
7. Функциональные возможности системы дыхания в художественной гимнастике
8. Особенности легочной вентиляции при занятиях художественной гимнастикой
Заключение
Литература
Введение
Многие годы человечество дышит и совершенно не замечает этого. При каждом вдохе воздух входит в легкие, и при каждом выдохе небольшая часть альвеолярного воздуха выходит в атмосферу. Однако именно этот отточенный до совершенства механизм есть крайне необходимая основа жизни человека.
Дыхание - это жизнь. Действительно, если без твердой пищи организм может обходиться несколько месяцев, без воды - несколько дней, то без воздуха - всего несколько минут.
Дыхание- синоним и неотъемлемый признак жизни."Пока дышу - надеюсь", утверждали древние римляне, а греки называли атмосферу "пастбищем жизни". Человек в день съедает примерно 1,24 кг пищи, выпивает 2 л воды, но вдыхает свыше 9 кг воздуха (более 10 000 л).
У людей, не занимающихся физическими упражнениями, спортом, разница в объеме грудной клетки при максимальном вдохе и выдохе не превышает обычно 4-б см, а у спортсменов она достигает 8-12 см и более. Дыхание может быть глубоким или поверхностным, редким или частым, правильным или неправильным. Правильным дыханием считается ритмичное глубокое дыхание, сопровождающееся полным расширением грудной клетки. Ритм дыхания может изменяться по разным причинам: от физического усилия, под влиянием температуры, при заболевании. По частоте дыхания можно судить и о влиянии физических упражнений на организм человека.
Усиление работы органов дыхания выражается в увеличении частоты и глубины дыхания, что значительно повышает легочную вентиляцию, т. е. увеличивается количество вдыхаемого и выдыхаемого воздуха.
Известно, что в покое легочная вентиляция у спортсменов равна 6-8 л в минуту, а при выполнении спортивных нагрузок (бег, ходьба на лыжах, плавание, езда на велосипеде) легочная вентиляция повышается до 120-130 л и более в минуту.
1. Характеристика понятия "дыхание"
Дыхание- это совокупность процессов, обеспечивающих потребление организмом кислорода и выделение двуокиси углерода. - В условиях покоя в организме за 1 минуту потребляется в среднем 250 - 300 мл О2 и выделяется 200 - 250 мл СО2. При физической работе большой мощности потребность в кислороде существенно возрастает и максимальное потребление кислорода (МПК) достигает у высокотренированных людей около 6 - 7 л/мин.
Дыхание осуществляет перенос О2 из атмосферного воздуха к тканям организма, а в обратном направлении производит удаление СО2 из организма в атмосферу.
Различают несколько этапов дыхания:
1. Внешнее дыхание - обмен газов между атмосферой и альвеолами.
Обмен газов между альвеолами и кровью легочных капилляров.
Транспорт газов кровью - процесс переноса О2 от легких к тканям и СО2 от тканей - к легким.
Обмен О2 и СО2 между кровью капилляров и клетками тканей организма.
Внутреннее, или тканевое, дыхание- биологическое окисление в митохондриях клетки.
Состав и свойства дыхательных сред
Дыхательной средой для человека является атмосферный воздух, состав которого отличается постоянством. В 1 л сухого воздуха содержится 780 мл азота, 210 мл кислорода и 0,3 мл двуокиси углерода (табл. 1). Остальные 10 мл приходятся на инертные газы - аргон, неон, гелий, криптон, ксенон и водород.
Таблица 1. Содержание и парциальное давление (напряжение) кислорода и углекислого газа в различных средах
СредаКислородУглекислый газ%мм рт. ст.мл/л%мм рт. ст.мл/лВдыхаемый воздух20,93159209,30,030,20,3Выдыхаемый воздух16,0121160,04,53445Альвеолярный воздух14,0100140,05,54055Артериальная кровь-100-96200,0-40560-540Венозная кровь-40140-160-46580Ткань-10-15--60-Около митохондрий-01-1--70-
На уровне моря нормальное атмосферное давление составляет 760 мм рт ст. Согласно закону Дальтона эта величина складывается из парциальных давлений всех газов, входящих в состав воздуха. Атмосферный воздух содержит также пары воды. В умеренном климате при температуре 22°С парциальное давление водяного пара в воздухе составляет 20 мм рт.ст. Парциальное давление водяного пара, уравновешенного в легких с кровью при атмосферном давлении 760 мм рт.ст. и температуре тела 37°С, составляет 47 мм рт.ст. Учитывая, что давление водяных паров в организме выше, чем в окружающей среде, в процессе дыхания организм теряет воду.
2. Внешнее дыхание
Внешнее дыхание осуществляется благодаря изменениям объема грудной клетки и сопутствующим изменениям объема легких. Во время вдоха объем грудной клетки увеличивается, а во время выдоха - уменьшается. В дыхательных движениях участвуют:
Дыхательные пути, которые по своим свойствам являются слегка растяжимыми, сжимаемыми и создают поток воздуха. Дыхательная система состоит из тканей и органов, обеспечивающих легочную вентиляцию и легочное дыхание (воздухоносные пути, легкие и элементы костно-мышечной системы). К воздухоносным путям, управляющим потоком воздуха, относятся: нос, полость носа, носоглотка, гортань, трахея, бронхи и бронхиолы. Легкие состоят из бронхиол и альвеолярных мешочков, а также из артерий, капилляров и вен легочного круга кровообращения. К элементам костно-мышечной системы, связанным с дыханием, относятся ребра, межреберные мышцы, диафрагма и вспомогательные дыхательные мышцы. Нос и полость носа служат проводящими каналами для воздуха, где он нагревается, увлажняется и фильтруется. Полость носа выстлана богато васкулиризированной слизистой оболочкой. В верхней части полости носа лежат обонятельные рецепторы. Носовые ходы открываются в носоглотку. Гортань лежит между трахеей и корнем языка. У нижнего конца гортани начинается трахея и спускается в грудную полость, где делится на правый и левый бронхи. Установлено, что дыхательные пути от трахеи до концевых дыхательных единиц (альвеол) ветвятся (раздваиваются) 23 раза. Первые 16 "поколений" дыхательных путей - бронхи и бронхиолы выполняют проводящую функцию. "Поколения" 17-22 - респираторные бронхиолы и альвеолярные ходы, составляют переходную (транзиторную) зону, и только 23-е "поколение" является дыхательной респираторной зоной и целиком состоит из альвеолярных мешочков с альвеолами. Общая площадь поперечного сечения дыхательных путей по мере ветвления возрастает более чем в 4,5 тысячи раз. Правый бронх обычно короче и шире левого.
Эластическая и растяжимая легочная ткань .Респираторный отдел представлен альвеолами. В легких имеется три типа альвеолоцитов (пневмоцитов), выполняющих разную функцию. Альвеолоциты второго типа осуществляют синтез липидов и фосфолипидов легочного сурфактанта. Общая площадь альвеол у взрослого человека достигает 80-90 м2, т.е. примерно в 50 раз превышает поверхность тела человека.
Грудная клетка , состоящая из пассивной костно-хрящевой основы, которая соединена соединительными связками и дыхательными мышцами, осуществляющими поднятие и опускание ребер и движения купола диафрагмы. За счет большого количества эластической ткани легкие, обладая значительной растяжимостью и эластичностью, пассивно следуют за всеми изменениями конфигурации и объема грудной клетки. Чем больше разность между давлением воздуха внутри и снаружи легкого, тем больше они будут растягиваться. Для иллюстрации этого положения служит модель Дондерса.
Рис.1. Модель Дондерса (а - экскурсия легких в конце выдоха; б - экскурсия легких во время вдоха)
Существуют два механизма, вызывающие изменение объема грудной клетки:поднятие и опускание реберидвижения купола диафрагмы. Дыхательные мышцы подразделяются наинспираторные и экспираторные .
Инспираторными мышцами являютсядиафрагма, наружные межреберные и межхрящевые мышцы. При спокойном дыхании объем грудной клетки изменяется в основном за счет сокращения диафрагмы и перемещения ее купола. всего на 1 см соответствует увеличение емкости грудной полости примерно на 200 - 300 мл. При глубоком форсированном дыхании участвуют дополнительные мышцы вдоха: трапециевидные, передние лестничные и грудино-ключично-сосцевидные мышцы. Они включаются в активный процесс дыхания при значительно больших величинах легочной вентиляции, например, при восхождении альпинистов на большие высоты или при дыхательной недостаточности, когда в процесс дыхания вступают почти все мышцы туловища.
Экспираторными мышцами являются внутренние межреберные и мышцы брюшной стенки, или мышцы живота. Каждое ребро способно вращаться вокруг оси, проходящей через две точки подвижного соединения с телом и поперечным отростком соответствующего позвонка.
Верхние отделы грудной клетки на вдохе расширяются преимущественно в переднезаднем направлении, а нижние отделы больше расширяются в боковых направлениях, так как ось вращения нижних ребер занимает сагиттальное положение.
В фазу вдоха наружные межреберные мышцы, сокращаясь, поднимают ребра, а в фазу выдоха ребра опускаются благодаря активности внутренних межреберных мышц.
При обычном спокойном дыхании выдох осуществляется пассивно, поскольку грудная клетка и легкие спадаются - стремятся занять после вдоха то положение, из которого они были выведены сокращением дыхательных мышц. Однако при кашле, рвоте, натуживании мышцы выдоха активны.
При спокойном вдохе увеличение объема грудной клетки составляет примерно 500-600 мл. Движение диафрагмы во время дыхания обусловливает до 80% вентиляции легких. У спортсменов высокой квалификации во время глубокого дыхания купол диафрагмы может смещаться до 10-12 см.
3. Легочная вентиляция
Величина легочной вентиляции определяется глубиной дыхания и частотой дыхательных движений.
Количественной характеристикой легочной вентиляции служит минутный объем дыхания (МОД) - объем воздуха, проходящий через легкие за 1 минуту. В покое частота дыхательных движений человека составляет примерно 16 в 1 минуту, а объем выдыхаемого воздуха - около 500 мл. Умножив частоту дыхания в 1 минуту на величину дыхательного объема, получим МОД, который у человека в покое составляет в среднем 8 л/мин.
Максимальная вентиляция легких(МВЛ) - объем воздуха, который проходит через легкие за 1 минуту во время максимальных по частоте и глубине дыхательных движений, Максимальная вентиляция возникает во время интенсивной работы, при недостатке содержания 02 (гипоксия) и избытке СО2 (гиперкапния) во вдыхаемом воздухе. В этих условиях МОД может достигать 150 - 200 л в 1 минуту.
Объем воздуха в легких и дыхательных путях зависит от конституционально-антропологических и возрастных характеристик человека, свойств легочной ткани, поверхностного натяжения альвеол, а также силы, развиваемой дыхательными мышцами.
Для оценки вентиляционной функции легких, состояния дыхательных путей, изучения паттерна (рисунка) дыхания применяются различные методы исследования:пневмография, спирометрия, спирография, пневмоскрин. С помощью спирографа можно определить и записать величины легочных объемов воздуха, проходящих через воздухоносные пути человека (рис. 2).
Рис.2. Легочные объемы и емкости (пояснения в тексте)
При спокойном вдохе и выдохе через легкие проходит сравнительно небольшой объем воздуха. Это дыхательный объем (ДО), который у взрослого человека составляет примерно 500 мл. При этом акт вдоха проходит несколько быстрее, чем акт выдоха. Обычно за 1 минуту совершается 12 - 16 дыхательных циклов. Такой тип дыхания обычно называется "эйпноэ" или "хорошее дыхание".
Прифорсированном (глубоком) вдохечеловек может дополнительно вдохнуть еще определенный объем воздуха. Этот резервный объем вдоха (РОвд) - максимальный объем воздуха, который способен вдохнуть человек после спокойного вдоха. Величина резервного объема вдоха составляет у взрослого человека примерно 1,8 - 2,0 л.
После спокойного выдоха человек может при форсированном выдохе дополнительно выдохнуть еще определенный объем воздуха. Это резервный объем выдоха (РОвыд), величина которого составляет в среднем 1,2 - 1,4 л.
Объем воздуха, который остается в легких после максимального выдоха и в легких мертвого человека, - остаточный объем легких (00). Величина остаточного объема составляет 1,2 - 1,5 л. У аборигенов высокогорья из-за бочкообразной грудной клетки сохраняются более высокие величины этого показателя, благодаря чему удается сохранить в организме необходимое содержание СО2, достаточное для регуляции дыхания в этих условиях.
общая емкость легких(ОЕЛ) - объем воздуха, находящегося в легких после максимального вдоха - все четыре объема;
жизненная емкость легких(ЖЕЛ) включает в себя дыхательный объем, резервный объем вдоха, резервный объем выдоха. ЖЕЛ - это объем воздуха, выдохнутого из легких после максимального вдоха при максимальном выдохе. ЖЕЛ = ОЕЛ - остаточный объем легких. ЖЕЛ составляет у мужчин 3,5 - 5,0 л, у женщин - 3,0-4,0л;
емкость вдоха(Ед.) равна сумме дыхательного объема и резервного объема вдоха, составляет в среднем 2,0 - 2,5 л;
функциональная остаточная емкость(ФОЕ) - объем воздуха в легких после спокойного выдоха. В легких при спокойном вдохе и выдохе постоянно содержится примерно 2500 мл воздуха, заполняющего альвеолы и нижние дыхательные пути. Благодаря этому газовый состав альвеолярного воздуха сохраняется на постоянном уровне.
Исследование легочных объемов и ёмкостей как важнейших показателей функционального состояния легких имеет большое медико-физиологическое значение не только для диагностики заболеваний (ателектаз, рубцовые изменения легких, поражения плевры), но и для экологического мониторинга местности и оценки состояния функции дыхания популяции в экологически неблагополучных зонах,
Для сопоставимости результатов измерений газовых объемов и емкостей материалы исследований должны быть приведены к стандартному состоянию BTPS, т.е. соотноситься с условиями в легких, где температура альвеолярного воздуха соответствует температуре тела, кроме того, воздух находится при определенном давлении и насыщен водяными парами.
Воздух, находящийся в воздухоносных путях (полость рта, носа, глотки, трахеи, бронхов и бронхиол), не участвует в газообмене, и поэтому пространство воздухоносных путей называют вредным или мертвым дыхательным пространством. Во время спокойного вдоха объемом 500 мл в альвеолы поступает только 350 мл вдыхаемого атмосферного воздуха. Остальные 150 мл задерживаются в анатомическом мертвом пространстве. Составляя в среднем треть дыхательного объема, мертвое пространство снижает на эту величину эффективность альвеолярной вентиляции при спокойном дыхании. В тех случаях, когда при выполнении физической работы дыхательный объем увеличивается в несколько раз, объем анатомического мертвого пространства практически не влияет на эффективность альвеолярной вентиляции.
При некоторых патологических состояниях - при анемии, легочной эмболии или эмфиземе могут возникать очаги - зоны альвеолярного мертвого пространства. В подобных зонах легких не происходит газообмена.
4. Обмен газов в легких и их перенос
Газообмен О2 и СО2 через альвеолярно-капиллярную мембрану происходит с помощью диффузии, которая осуществляется в два этапа. На первом этапе диффузионный перенос газов происходит через аэрогематический барьер, на втором - происходит связывание газов в крови легочных капилляров, объем которой оставляет 80-150 мл при толщине слоя крови в капиллярах всего 5-8 мкм. Плазма крови практически не препятствует диффузии газов, в отличие от мембраны эритроцитов.
Структура легких создает благоприятные условия для газообмена: дыхательная зона каждого легкого содержит около 300 млн. альвеол и примерно такое же число капилляров, имеет площадь 40-140 м2, при толщине аэрогематического барьера всего 0,3-1,2 мкм.
Особенности диффузии газов количественно характеризуются через диффузионную способность легких. Для О2 диффузионная способность легких - это объем газа, переносимого из альвеол в кровь в 1 минуту при градиенте альвеолярно-капиллярного давления газа, равном 1 мм рт.ст.
Движение газов происходит в результате разницы парциальных давлений. Парциальное давление - это та часть давления, которую составляет данный газ из общей смеси газов. Пониженное давление Од в ткани способствует движению кислорода к ней. Для СО2 градиент давления направлен в обратную сторону, и СО2 с выдыхаемым воздухом уходит в окружающую среду. Изучение физиологии дыхания фактически сводится к изучению этих градиентов и того, как они поддерживаются.
Градиент парциального давления кислорода и углекислого газа это сила, с которой молекулы этих газов стремятся проникнуть через альвеолярную мембрану в кровь. Парциальное напряжение газа в крови или тканях - это сила, с которой молекулы растворимого газа стремятся выйти в газовую среду.
На уровне моря атмосферное давление составляет в среднем 760 мм рт.ст., а процентное содержание кислорода - около 21%. В этом случае рО2 в атмосфере составляет: 760 х 21/100=159 мм рт.ст. При вычислении парциального давления газов в альвеолярном воздухе следует учитывать, что в этом воздухе присутствуют пары воды (47 мм рт.ст.). Поэтому это число вычитают из значения атмосферного давления, и на долю парциального давления газов приходится (760^47) =713 мм рт.ст. При содержании кислорода в альвеолярном воздухе, равном 14 %, его парциальное давление будет 100 мм рт. ст. При содержании двуокиси углерода, равном 5,5%, парциальное давление СО2 составит примерно 40 мм рт.ст.
В артериальной кровипарциальное напряжение кислородадостигает почти 100 мм рт.ст., в венозной крови - около 40 мм рт.ст., а в тканевой жидкости, в клетках - 10-15 мм рт.ст. Напряжение углекислого газа в артериальной крови составляет около 40 мм рт.ст., в венозной - 46 мм рт.ст., а в тканях - до 60 мм рт.ст.
Газы в крови находятся в двух состояниях: физически растворенном и химически связанном. Растворение происходит в соответствии с законом Генри, согласно которому количество газа, растворенного в жидкости, прямо пропорционально парциальному давлению этого газа над жидкостью. На каждую единицу парциального давления в 100 мл крови растворяется 0,003 мл О2 или 3 мл/л крови.
Каждый газ имеет свойкоэффициент растворимости. При температуре тела растворимость СО2 в 25 раз больше, чем О2 Из-за хорошей растворимости углекислоты в крови и тканях СО2 переносится в 20 раз легче, чем О2 Стремление газа переходить из жидкости в газовую фазу называют напряжением газа. В обычных условиях в 100 мл крови находится в растворенном состоянии всего 0,3 мл 02 и 2,6 мл СО2 Такие величины не могут обеспечить запросы организма в О2
Газообмен кислорода между альвеолярным воздухом и кровью происходит благодаря наличию концентрационного градиента 02 между этими средами. Транспорт кислорода начинается в капиллярах легких, где основная масса поступающего в кровь О2 вступает в химическую связь с гемоглобином. Гемоглобин способен избирательно связывать 02 и образовывать оксигемоглобин (НвО2). Один грамм гемоглобина связывает 1,36 - 1,34 мл О2 а в 1 литре крови содержится 140-150 г гемоглобина. На 1 грамм гемоглобина приходится 1,39 мл кислорода. Следовательно, в каждом литре крови максимально возможное содержание кислорода в химически связанной форме составит 190 - 200 мл О2 или 19 об% - это кислородная емкость крови. Кровь человека содержит примерно 700 - 800 г гемоглобина и может связывать 1 л кислорода.
Подкислородной емкостью кровипонимают количество О2 которое связывается кровью до полного насыщения гемоглобина. Изменение концентрации гемоглобина в крови, например, при анемиях, отравлениях ядами изменяет ее кислородную емкость. При рождении в крови у человека более высокие значения кислородной емкости и концентрации гемоглобина. Насыщение крови кислородом выражает отношение количества связанного кислорода к кислородной емкости крови, т.е. под насыщением крови 02 подразумевается процент оксигемоглобина по отношению к имеющемуся в крови гемоглобину. В обычных условиях насыщение О2 составляет 95-97%. При дыхании чистым кислородом насыщение крови 02 достигает 100%, а при дыхании газовой смесью с низким содержанием кислорода процент насыщения падает. При 60-65% наступает потеря сознания.
Зависимость связывания кислорода кровью от его парциального давления можно представить в виде графика, где по оси абсцисс откладывается р02 в крови, по ординате - насыщение гемоглобина кислородом. Этот график - кривая диссоциации оксигемоглобина, или сатурационная кривая, показывает, какая доля гемоглобина в данной крови связана с 02 при том или ином его парциальном давлении, а какая - диссоциирована, т.е. свободна от Кислорода. Кривая диссоциации имеет S-образную форму. Плато кривой характерно для насыщенной О2 (сатурированной) артериальной крови, а крутая нисходящая часть кривой - венозной, или десатурированной, крови в тканях (рис. 3).
Рис. 3. Кривые диссоциации оксигемоглобина цельной крови при различных рН крови [А] и при изменении температуры (5)
Кривые 1-6 соответствуют 0°, 10°, 20°, 30°, 38° и 43°С
Сродство кислорода к гемоглобину и способность отдавать 02 в тканях зависит от метаболических потребностей клеток организма и регулируется важнейшими факторами метаболизма тканей, вызывающими смещение кривой диссоциации. К этим факторам относятся: концентрация водородных ионов, температура, парциальное напряжение углекислоты и соединение, которое накапливается в эритроцитах - это 2,3-дифосфоглицератфосфат (ДФГ). Уменьшение рН крови вызывает сдвиг кривой диссоциации вправо, а увеличение рН крови - сдвиг кривой влево. Вследствие повышенного содержания СО2 в тканях рН также меньше, чем в плазме крови. Величина рН и содержание СО2 в тканях организма изменяют сродство гемоглобина к О2. Их влияние на кривую диссоциации оксигемоглобина называется эффектом Бора (Х. Бор, 1904). При повышении концентрации водородных ионов и парциального напряжения СО2 в среде сродство гемоглобина к кислороду снижается. Этот "эффект" имеет важное приспособительное значение: СО2 в тканях поступает в капилляры, поэтому кровь при том же рО2 способна освободить больше кислорода. Образующийся при расщеплении глюкозы метаболит 2,3-ДФГ также снижает сродство гемоглобина к кислороду.
На кривую диссоциации оксигемоглобина оказывает влияние также и температура. Рост температуры значительно увеличивает скорость распада оксигемоглобина и уменьшает сродство гемоглобина к 02. Увеличение температуры в работающих мышцах способствует освобождению О2 Связывание 02 гемоглобином снижает сродство его аминогрупп к СО2 (эффект Холдена). Диффузия СО2 из крови в альвеолы обеспечивается за счет поступления растворенного в плазме крови СО2 (5-10%), из гидрокарбонатов (80-90%) и, наконец, из карбаминовых соединений эритроцитов (5-15%), которые способны диссоциировать.
Углекислый газ в крови находится в трех фракциях: физически растворенный, химически связанный в виде бикарбонатов и химически связанный с гемоглобином в виде карбогемоглобина. В венозной крови углекислого газа содержится всего 580 мл. При этом на долю физически растворенного газа приходится 25 мл, на долю карбогемоглобина - около 45 мл, на долю бикарбонатов - 510 мл (бикарбонатов плазмы - 340 мл, эритроцитов - 170 мл). В артериальной крови содержание угольной кислоты меньше.
От парциального напряжения физически растворенного углекислого газа зависит процесс связывания СО2 кровью. Углекислота поступает в эритроцит, где имеется фермент карбоангидраза, который может в 10 000 раз увеличить скорость образования угольной кислоты. Пройдя через эритроцит, угольная кислота превращается в бикарбонат и переносится к легким.
Эритроциты переносят в 3 раза больше СО2 чем плазма. Белки плазмы составляют 8 г на 100 см3 крови, гемоглобина же содержится в крови 15 г на 100 см3. Большая часть СО2 транспортируется в организме в связанном состоянии в виде гидрокарбонатов и карбаминовых соединений, что увеличивает время обмена СО2.
Кроме физически растворенного в плазме крови молекулярного СО2 из крови в альвеолы легких диффундирует СО2 который высвобождается из карбаминовых соединений эритроцитов благодаря реакции окисления гемоглобина в капиллярах легкого, а также из гидрокарбонатов плазмы крови в результате их быстрой диссоциации с помощью содержащегося в эритроцитах фермента карбоангидразы. Этот фермент в плазме отсутствует. Бикарбонаты плазмы для освобождения СО2 должны сначала проникнуть в эритроциты, чтобы подвергнуться действию карбоангидразы. В плазме находится бикарбонат натрия, а в эритроцитах - бикарбонат калия. Мембрана эритроцитов хорошо проницаема для СО2 поэтому часть СО2 быстро диффундирует из плазмы внутрь эритроцитов. Наибольшее количество бикарбонатов плазмы крови образуется при участии карбоангидразы эритроцитов.
Следует отметить, что процесс выведения СО2 из крови в альвеолы легкого менее лимитирован, чем оксигенация крови, так как молекулярный СО2 легче проникает через биологические мембраны, чем О2.
Различные яды, ограничивающие транспорт Од, такие как СО, нитриты, ферроцианиды и многие другие, практически не действуют на транспорт СО2 Блокаторы карбоангидразы также никогда полностью не нарушают образование молекулярного СО2. И наконец, ткани обладают большой буферной емкостью, но не защищены от дефицита О2. Выведение СО2 легкими может нарушиться при значительном уменьшении легочной вентиляции (гиповентиляции) в результате заболевания легких, дыхательных путей, интоксикации или нарушении регуляции дыхания. Задержка СО2 приводит к дыхательному ацидозу - уменьшению концентрации бикарбонатов, сдвигу рН крови в кислую сторону. Избыточное выведение СО2 при гипервентиляции во время интенсивной мышечной работы, при восхождении на большие высоты может вызвать дыхательный алкалоз, сдвиг рН крови в щелочную сторону.
5. Регуляция дыхания
Регуляция внешнего дыхания. В соответствии с метаболическими потребностями дыхательная система обеспечивает газообмен О2 и СО2 между окружающей средой и организмом. Эту жизненно важную функцию регулирует сеть многочисленных взаимосвязанных нейронов ЦНС, расположенных в нескольких отделах мозга и объединяемых в комплексное понятие"дыхательный центр". При воздействии на его структуры нервных и гуморальных стимулов происходит приспособление функции дыхания к меняющимся условиям внешней среды. Структуры, необходимые для возникновения дыхательного ритма, впервые были обнаружены в продолговатом мозге. Перерезка продолговатого мозга в области дна IV желудочка приводит к прекращению дыхания. Поэтому под главным дыхательным центром понимают совокупность нейронов специфических дыхательных ядер продолговатого мозга.
Дыхательный центруправляет двумя основными функциями: двигательной, которая проявляется в виде сокращения дыхательных мышц, и гомеостатической, связанной с поддержанием постоянства внутренней среды организма при сдвигах в ней содержания 02 и СО2. Двигательная , или моторная, функция дыхательного центра заключается в генерации дыхательного ритма и его паттерна. Благодаря этой функции осуществляется интеграция дыхания с другими функциями. Под паттерном дыхания следует иметь в виду длительность вдоха и выдоха, величину дыхательного объема, минутного объема дыхания. Гомеостатическая функция дыхательного центра поддерживает стабильные величины дыхательных газов в крови и внеклеточной жидкости мозга, адаптирует дыхательную функцию к условиям измененной газовой среды и другим факторам среды обитания.
Локализация и функциональные свойства дыхательных нейронов.
В передних рогах спинного мозга на уровне С3 - С5 располагаются мотонейроны, образующие диафрагмальный нерв. Мотонейроны, иннервирующие межреберные мышцы, находятся в передних рогах на уровнях Т2 - Т10 (Т2 - Т6 - мотонейроны инспираторных мышц, T8-T10 - экспираторных). Установлено, что одни мотонейроны регулируют преимущественно дыхательную, а другие - преимущественно познотоническую активность межреберных мышц.
Нейроны бульбарного дыхательного центра располагаются на дне IV желудочка в медиальной части ретикулярной формации продолговатого мозга и образуют дорсальную и вентральную дыхательные группы. Дыхательные нейроны, активность которых вызывает инспирацию или экспирацию, называются соответственно инспираторными и экспираторными нейронами. Между группами нейронов, управляющими вдохом и выдохом, существуют реципрокные отношения. Возбуждение экспираторного центра сопровождается торможением в инспираторном центре и наоборот. Инспираторные и экспираторные нейроны, в свою очередь, делятся на "ранние" и "поздние". Каждый дыхательный цикл начинается с активизации "ранних" инспираторных нейронов, затем возбуждаются "поздние" инспираторные нейроны. Также последовательно возбуждаются "ранние" и "поздние" экспираторные нейроны, которые тормозят инспираторные нейроны и прекращают вдох. Современные исследования показали, что в продолговатом мозге нет четкого деления на инспираторный и экспираторный отделы, а есть скопления дыхательных нейронов с определенной функцией.
Спонтанная активность нейронов дыхательного центра начинает появляться к концу периода внутриутробного развития. Возбуждение дыхательного центра у плода появляется благодаря пейсмекерным свойствам сети дыхательных нейронов продолговатого мозга. По мере формирования синаптических связей дыхательного центра с различными отделами ЦНС пейсмекерный механизм дыхательной активности постепенно теряет свое физиологическое значение.
В варолиевом мосту находятся ядра дыхательных нейронов, образующих пневмотаксический центр. Считается, что дыхательные нейроны моста участвуют в механизме смены вдоха и выдоха и регулируют величину дыхательного объема. Дыхательные нейроны продолговатого мозга и варолиева моста связаны между собой восходящими и нисходящими нервными путями и функционируют согласованно. Получив импульсы от инспираторного центра продолговатого мозга, пневмотаксический центр посылает их к экспираторному центру продолговатого мозга, возбуждая последний. Инспираторные нейроны тормозятся. Разрушение мозга между продолговатым мозгом и мостом удлиняет фазу вдоха. Гипоталамические ядра координируют связь дыхания с кровообращением.
Определенные зоны коры больших полушарий осуществляют произвольную регуляцию дыхания в соответствии с особенностями влияния на организм факторов внешней среды и связанными с этим гомеостатическими сдвигами.
Таким образом, мы видим, что управление дыханием - сложнейший процесс, осуществляемый множеством нейронных структур. В процессе управления дыханием осуществляется четкая иерархия различных компонентов и структур дыхательного центра.
Рефлекторная регуляция дыхания.
Нейроны дыхательного центра имеют связи с многочисленными механорецепторами дыхательных путей и альвеол легких и рецепторов сосудистых рефлексогенных зон. Благодаря этим связям осуществляется весьма многообразная, сложная и биологически важная рефлекторная регуляция дыхания и ее координация с другими функциями организма.
Различают несколько типов механорецепторов: медленно адаптирующиеся рецепторы растяжения легких, ирритантные быстро адаптирующиеся механорецепторы и J-рецепторы - "юкстакапиллярные" рецепторы легких.
Медленно адаптирующиеся рецепторы растяжения легкихрасположены в гладких мышцах трахеи и бронхов. Эти рецепторы возбуждаются при вдохе, импульсы от них по афферентным волокнам блуждающего нерва поступают в дыхательный центр. Под их влиянием тормозится активность инспираторных нейронов продолговатого мозга. Вдох прекращается, начинается выдох, при котором рецепторы растяжения неактивны. Рефлекс торможения вдоха при растяжении легких называется рефлексом Геринга - Брейера. Этот рефлекс контролирует глубину и частоту дыхания. Он является примером регуляции по принципу обратной связи. После перерезки блуждающих нервов дыхание становится редким и глубоким.
Ирритантные быстро адаптирующиеся механорецепторы, локализованные в слизистой оболочке трахеи и бронхов, возбуждаются при резких изменениях объема легких, при растяжении или спадении легких, при действии на слизистую трахеи и бронхов механических или химических раздражителей. Результатом раздражения ирритантных рецепторов является частое, поверхностное дыхание, кашлевой рефлекс, или рефлекс бронхоконстрикции.рецепторы - "юкстакапиллярные" рецепторы легких находятся в интерстиции альвеол и дыхательных бронхов вблизи от капилляров. Импульсы от J-рецепторов при повышении давления в малом круге кровообращения, или увеличении объема интерстициальной жидкости в легких (отек легких), или эмболии мелких легочных сосудов, а также при действии биологически активных веществ (никотин, простагландины, гистамин) по медленным волокнам блуждающего нерва поступают в дыхательный центр - дыхание становится частым и поверхностным (одышка).
Важное биологическое значение, особенно в связи с ухудшением экологических условий и загрязнением атмосферы, имеютзащитные дыхательные рефлексы - чихание и кашель.
Чихание. Раздражение рецепторов слизистой оболочки полости носа, например, пылевыми частицами или газообразными наркотическими веществами, табачным дымом, водой вызывает сужение бронхов, брадикардию, снижение сердечного выброса, сужение просвета сосудов кожи и мышц. Различные механические и химические раздражения слизистой оболочки носа вызывают глубокий сильный выдох - чихание, способствующее стремлению избавиться от раздражителя. Афферентным путем этого рефлекса является тройничный нерв.
Кашель возникает при раздражении механо- и хеморецепторов глотки, гортани, трахеи и бронхов. При этом после вдоха сильно сокращаются мышцы выдоха, резко повышается внутригрудное и внутрилегочное давление (до 200 мм рт. ст.), открывается голосовая щель, и воздух из дыхательных путей под большим напором высвобождается наружу и удаляет раздражающий агент. Кашлевой рефлекс является основным легочным рефлексом блуждающего нерва.
Рефлексы с проприорецепторов дыхательных мышц.
От мышечных веретен и сухожильных рецепторов Гольджи, расположенных в межреберных мышцах и мышцах живота, импульсы поступают в соответствующие сегменты спинного мозга, затем в продолговатый мозг, центры головного мозга, контролирующие состояние скелетных мышц. В результате происходит регуляция силы сокращений в зависимости от исходной длины мышц и оказываемого им сопротивления дыхательной системы.
Рефлекторная регуляция дыхания осуществляется также периферическими и центральными хеморецепторами, что изложено в разделе гуморальной регуляции.
Гуморальная регуляция дыхания.
Главным физиологическим стимулом дыхательных центров является двуокись углерода. Регуляция дыхания обусловливает поддержание нормального содержания СО2 в альвеолярном воздухе и артериальной крови. Возрастание содержания СО2 в альвеолярном воздухе на 0,17% вызывает удвоение МОД, а вот снижение О2 на 39-40% не вызывает существенных изменений МОД.
При повышении в замкнутых герметических кабинах концентрации СО2 до 5 - 8% у обследуемых наблюдалось увеличение легочной вентиляции в 7-8 раз. При этом концентрация СО2 в альвеолярном воздухе существенно не возрастала, так как основным признаком регуляции дыхания является необходимость регуляции объема легочной вентиляции, поддерживающей постоянство состава альвеолярного воздуха.
Деятельность дыхательного центра зависит от состава крови, поступающей в мозг по общим сонным артериям. В 1890 г. это было показано Фредериком в опытах с перекрестным кровообращением. У двух собак, находившихся под наркозом, перерезали и соединяли перекрестно сонные артерии и яремные вены. При этом голова первой собаки снабжалась кровью второй собаки и наоборот. Если у одной из собак, например у первой, перекрывали трахею и таким путем вызывали асфиксию, то гиперпноэ развивалось у второй собаки. У первой же собаки, несмотря на увеличение в артериальной крови напряжения СО2 и снижение напряжения 02, развивалось апноэ, так как в ее сонную артерию прступала кровь второй собаки, у которой в результате гипервентиляции снижалось напряжение СО2 в артериальной крови.
Двуокись углерода, водородные ионы и умеренная гипоксия вызывают усиление дыхания. Эти факторы усиливают деятельность дыхательного центра, оказывая влияние на периферические (артериальные) и центральные (модулярные) хеморецепторы, регулирующие дыхание.
Артериальные хеморецепторы находятся в каротидных синусах и дуге аорты. Они расположены в специальных тельцах, обильно снабжаемых артериальной кровью. Аортальные хеморецепторы на дыхание влияют слабо и большее значение имеют для регуляции кровообращения.
Артериальные хеморецепторы являются уникальными рецепторными образованиями, на которые гипоксия оказывает стимулирующее влияние. Афферентные влияния каротидных телец усиливаются также при повышении в артериальной крови напряжения двуокиси углерода и концентрации водородных ионов. Стимулирующее действие гипоксии и гиперкапнии на хеморецепторы взаимно усиливается, тогда как в условиях гипероксии чувствительность хеморецепторов к двуокиси углерода резко снижается. Артериальные хеморецепторы информируют дыхательный центр о напряжении 02 и СО2 в крови, направляющейся к мозгу.
После перерезки артериальных (периферических) хеморецепторов у подопытных животных исчезает чувствительность дыхательного центра к гипоксии, но полностью сохраняется реакция дыхания на гиперкапнию и ацидоз.
Центральные хеморецепторы расположены в продолговатом мозге латеральнее пирамид. Перфузия этой области мозга раствором со сниженным рН резко усиливает дыхание, а при высоком рН дыхание ослабевает, вплоть до апноэ. То же происходит при охлаждении или обработке этой поверхности продолговатого мозга анестетиками. Центральные хеморецепторы, оказывая сильное влияние на деятельность дыхательного центра, существенно изменяют вентиляцию легких. Установлено, что снижение рН спинномозговой жидкости всего на 0,01 сопровождается увеличением легочной вентиляции на 4 л/мин.
Центральные хеморецепторы реагируют на изменение напряжения СО2 в артериальной крови позже, чем периферические хеморецепторы, так как для диффузии СО2 из крови в спинномозговую жидкость и далее в ткань мозга необходимо больше времени. Гиперкапния и ацидоз стимулируют, а гипокапния и алкалоз - тормозят центральные хеморецепторы.
Для определения чувствительности центральных хеморецепторов к изменению рН внеклеточной жидкости мозга, изучения синергизма и антагонизма дыхательных газов, взаимодействия системы дыхания и сердечно-сосудистой системы используют метод возвратного дыхания. При дыхании в замкнутой системе выдыхаемый СОд вызывает линейное увеличение концентрации СО2 и одновременно повышается концентрация водородных ионов в крови, а также во внеклеточной жидкости мозга.
Совокупность дыхательных нейронов следовало бы рассматривать как созвездие структур, осуществляющих центральный механизм дыхания. Таким образом, вместо термина "дыхательный центр" правильнее говорить о системе центральной регуляции дыхания, которая включает в себя структуры коры головного мозга, определенные зоны и ядра промежуточного, среднего, продолговатого мозга, варолиева моста, нейроны шейного и грудного отделов спинного мозга, центральные и периферические хеморецепторы, а также механорецепторы органов дыхания.
Своеобразие функции внешнего дыхания состоит в том, что она одновременно и автоматическая, и произвольно управляемая.
. Особенности дыхания при физической нагрузке и при измененном парциальном давлении.
В различных условиях среды обитания системы нейрогуморальной регуляции дыхания и кровообращения функционируют в тесном взаимодействии как единая кардиореспираторная система. Особенно четко это проявляется при интенсивной физической нагрузке и в условиях гипоксии - недостаточном снабжении организма кислородом. В процессе жизнедеятельности в организме возникают различные виды гипоксии, имеющие эндогенную и экзогенную природу.
Во время выполнения физической работы мышцам необходимо большое количество кислорода. Потребление 02 и продукция СО2 возрастают при физической нагрузке в среднем в 15 - 20 раз. Обеспечение организма кислородом достигается сочетанным усилением функции дыхания и кровообращения. Уже в начале мышечной работы вентиляция легких быстро увеличивается. В возникновении гиперпноэ в начале физической работы периферические и центральные хеморецепторы как важнейшие чувствительные структуры дыхательного центра еще не участвуют. Уровень вентиляции в этот период регулируется сигналами, поступающими к дыхательному центру главным образом из гипоталамуса, лимбической системы и двигательной зоны коры большого мозга, а также раздражением проприорецепторов работающих мышц. По мере продолжения работы к нейрогенным стимулам присоединяются гуморальные воздействия, вызывающие дополнительный прирост вентиляции. При тяжелой физической работе на уровень вентиляции оказывают влияние также повышение температуры, артериальная двигательная гипоксия и другие лимитирующие факторы.
Таким образом, наблюдаемые при физической работе изменения дыхания обеспечиваются сложным комплексом нервных и гуморальных механизмов. Однако из-за индивидуально лимитирующих факторов биомеханики дыхания, особенностей экопортрета человека, не всегда удается при выполнении одной и той же нагрузки полностью объяснить точное соответствие вентиляции легких уровню метаболизма в мышцах.
Экзогенная гипоксия развивается в результате действия измененных (в сравнении с обычными) факторов внешней среды.
Эндогенная гипоксия возникает при различных физиологических и патологических изменениях в различных функциональных системах организма.
Реакция внешнего дыхания на гипоксию зависит от продолжительности и скорости нарастания гипоксического воздействия, степени потребления кислорода (покой и физическая нагрузка), индивидуальных особенностей организма и совокупности генетически обусловленных свойств и наследственных морфофункциональных признаков (экопортрет коренных жителей высокогорья и популяции различных этнических групп).
Наблюдаемая в условиях кислородной недостаточности первоначальная гипоксическая стимуляция дыхания приводит к вымыванию углекислоты из крови и развитию дыхательного алкалоза. Гипоксия сочетается с гипокапнией. В свою очередь, это способствует увеличению рН внеклеточной жидкости мозга. Центральные хеморецепторы реагируют на подобный сдвиг рН в цереброспинальной жидкости мозга резким снижением своей активности. Это вызывает настолько существенное торможение нейронов дыхательного центра, что он становится нечувствительным к стимулам, исходящим от периферических хеморецепторов. Наступает своеобразная гипоксическая "глухота". Несмотря на сохраняющуюся гипоксию, постепенно гиперпноэ сменяется непроизвольной гиповентиляцией, что в определенной мере способствует также сохранению физиологически необходимого количества углекислоты.
Реакция на гипоксию у коренных жителей высокогорья и у горных животных практически отсутствует, и, по мнению многих авторов, у жителей равнин гипоксическая реакция также исчезает после продолжительной (не менее 3-5 лет) их адаптации к условиям высокогорья.
Основными факторами долговременной акклиматизации к условиям высокогорья являются; повышение содержания углекислоты и понижение содержания кислорода в крови на фоне снижения чувствительности периферических хеморецепторов к гипоксии, увеличения плотности капилляров и относительно высокого уровня утилизации тканями 02 из крови. У горцев также возрастают диффузионная способность легких и кислородная емкость крови за счет роста концентрации гемоглобина. Одним из механизмов, позволяющих горцам в условиях гипоксии повысить отдачу кислорода тканям и сохранить углекислоту, является способность повышенного образования у них метаболита глюкозы - 2,3 дифосфоглицерата. Этот метаболит снижает сродство гемоглобина к кислороду.
Предметом интенсивных физиологических исследований как в эксперименте, так и в различных природно-климатических и производственных условиях является изучение функционального взаимодействия систем регуляции дыхания и кровообращения. Обе системы имеют общие рефлексогенные зоны в сосудах, которые посылают афферентные сигналы к специализированным нейронам основного чувствительного ядра продолговатого мозга - ядра одиночного пучка. Здесь же в непосредственной близости находятся дорсальное ядро дыхательного центра и сосудодвигательный центр. Особо следует отметить, что легкие являются единственным органом, куда поступает весь минутный объем крови. Это обеспечивает не только газотранспортную функцию, но и роль своеобразного фильтра, который определяет состав биологически активных веществ в крови и их метаболизм.
Дыхание при высоком атмосферном давлении.
Во время водолазных и кессонных работ человек находится под давлением выше атмосферного на 1 атм. на каждые 10 м погружения. В этих условиях увеличивается количество газов, растворенных в крови, и особенно азота. При быстром подъеме водолаза на поверхность физически растворенные в крови и тканях газы не успевают выделиться из организма и образуют пузырьки - кровь "закипает". Кислород и углекислый газ быстро связываются кровью и тканями. Особую опасность представляют пузырьки азота, которые разносятся кровью и закупоривают мелкие сосуды (газовая эмболия), что сопровождается тяжелыми повреждениями ЦНС, органов зрения, слуха, сильными болями в мышцах и в области суставов, потерей сознания. Такое состояние, возникающее при быстрой декомпрессии, называется кессонной болезнью. Пострадавшего необходимо вновь поместить в среду с высоким давлением, а затем постепенно производить декомпрессию.
Вероятность возникновения кессонной болезни может быть значительно снижена при дыхании специальными газовыми смесями, например гелиево-кислородной. Гелий почти нерастворим в крови, он быстрее диффундирует из тканей.
. Функциональные возможности системы дыхания в художественной гимнастике
Высокие требования к ловкости и гибкости спортсмена предъявляют сложнокоординационные виды спорта, каким являются акробатика, аэробика, гимнастика спортивная и художественная, прыжки в воду и на батуте, прыжки на лыжах с трамплина, слалом, фристайл, фигурное катание и др.Занятия, особенно гимнастическими и акробатическими упражнениями, оказывают мощное стимулирующее воздействие на опорно-двигательный аппарат. Тренировочный эффект выражается в существенном увеличении подвижности в суставах при одновременном укреплении связочного аппарата, возрастании силовых возможностей мышц в динамических и статических нагрузках, повышении эластичности мышечных тканей. Все эти качества позволяют производить движения с большой амплитудой и высокой скоростью.
Для художественной гимнастики характерны особенно высокие требования к дифференцированию пространственно-временных и силовых показателей при действиях спортсменок со снарядами в условиях ограниченного визуального контроля. Большой акцент на увеличение амплитуды движений в художественной гимнастике выдвигает на первый план развитие такого качества, как гибкость, и снижает значимость силовой подготовки, в то время как в спортивной гимнастике и акробатике основные проблемы тренировки связаны с воспитанием силовых качеств. Надо отметить также присущую данной группе видов спорта эстетическую направленность.
В художественной гимнастике спортсменки выполняют свою программу под музыкальное сопровождение. Известно, что звучание музыкальных произведений повышает работоспособность сердечно-сосудистой, мышечной, дыхательной систем организма. При выполнении упражнений с музыкальным сопровождением улучшается легочная вентиляция, увеличивается амплитуда дыхательных движений.В то же время можно говорить о развитии у детей музыкальности, основных ее компонентов - эмоциональной отзывчивости, слуха. Ребенок и здесь учится воспринимать музыку, двигаться в соответствии с ее характером, средствами выразительности.
8. Особенности легочной вентиляции при занятиях художественной гимнастикой
дыхание гомеостатический кровь газ
При занятиях художественной гимнастикой дыхание спортсменки становится совершеннее, а, следовательно, улучшаются и окислительные процессы, столь важные для всех жизненных функций. Легочная вентиляция увеличивается, частота дыханий уменьшается, что дает экономию в работе дыхательной мускулатуры, которая становится более сильной и выносливой. Подвижность грудной клетки и диафрагмы возрастает. Более совершенное дыхание благоприятно влияет и на процесс кровообращения. Регулярные и правильно проводимые занятия спортом развивают и улучшают функциональную способность аппарата дыхания. Дыхание становится глубоким, уреженным, повышается жизненная емкость легких.
У хорошо тренированного спортсмена и потребление кислорода, и общая вентиляция легких увеличиваются примерно в 20 раз при изменении интенсивности физической нагрузки от состояния покоя до максимального уровня. Максимальные возможности дыхательной системы примерно на 50% выше, чем истинное усиление дыхания во время максимальной мышечной работы. Это создает элемент надежности для спортсменов, обеспечивая им дополнительную вентиляцию, которая может стать необходимой при условиях: 1) мышечной работы на большой высоте; 2) физической работы в условиях очень высокой температуры; 3) патологии дыхательной системы.
Диффузионная способность кислорода у спортсменов . Диффузионная способность кислорода является мерой скорости диффузии кислорода из легочных альвеол в кровь. Величина этого показателя выражается в миллилитрах кислорода, способного диффундировать в 1 мин при разности парциального давления кислорода между альвеолами и кровью в легких, равной 1 мм рт. ст. Следовательно, если парциальное давление кислорода в альвеолах равно 91 мм рт. ст., а давление кислорода в крови составляет 90 мм рт. ст., количество кислорода, которое диффундирует через респираторную мембрану каждую минуту, равно его диффузионной способности. Далее приведены величины разной диффузионной способности.
Самое удивительное в этих результатах - увеличение в несколько раз диффузионной способности в состоянии максимальной физической нагрузки по сравнению с состоянием покоя. Это связано главным образом с тем, что в покое во многих легочных капиллярах кровоток снижен или даже практически отсутствует, тогда как при максимальной мышечной нагрузке увеличение легочного кровотока ведет к максимальной скорости перфузии всех легочных капилляров, что обеспечивает гораздо большую площадь поверхности, через которую кислород может диффундировать в кровь.
Спортсмены с большей минутной потребностью в кислороде имеют более высокую диффузионную способность.
Заключение
В организме человека нет запасов кислорода, поэтому его непрерывное поступление жизненно необходимо. Прекращение доступа кислорода в клетки организма приводит к их гибели. Образующийся при окислении веществ углекислый газ должен быть удален из организма, так как его накопление в значительном количестве опасно для жизни. Обмен кислорода и углекислого газа между организмом и окружающей средой называется дыханием .
Жизнь человека невозможна без дыхания, без поглощения кислорода воздуха и выделения образовавшейся в организме углекислоты. Работа аппарата дыхания, как и деятельность сердца, происходит в течение всей жизни человека непрерывно.
Человек и все высокоорганизованные живые существа нуждаются для своей нормальной жизнедеятельности в постоянном поступлении к тканям организма кислорода, который используется в сложном биохимическом процессе окисления питательных веществ, в результате чего выделяется энергия и образуется двуокись углерода и вода.
Занятия спортом и физическая работа повышают потребление кислорода работающими мышцами. В связи с этим деятельность органов дыхания усиливается, то же самое наблюдается в кровообращении, обмене веществ и др. Занятия спортом хорошо развивают и укрепляют органы дыхания; под их влиянием увеличиваются размеры грудной клетки, она приобретает красивую выпуклую форму, подвижность ее резко возрастает.
Литература
1.Агаджанян Н.А.,Шабатура Н.Н. Биоритмы, спорт, здоровье. - М.: ФиС, 1989.
2.Власова З.А.Биология. Справочник абитуриента - М.: Филол. об-во "Слово", АСТ ИД "Ключ-С", Центр гуманитарных наук при факультете журналистики МГУ им. М.В. Ломоносова, 1997.
Волков В.М.Тренеру о подростке. - М.: ФиС, 1973.
Волков Л.В.Физическое воспитание учащихся: Учебно-методическое пособие. - Киев: Рад. шк., 1988.
Кассиль Г.Н.Внутренняя среда организма. Изд. 2-е доп. и перераб. - М.: Наука, 1983.
Липченко В.Я., Самусев Р.П.Атлас нормальной анатомии человека. - М.: Медицина, 1984.
Полтырев С.С., Русин В.Я.Внутренние органы при физических нагрузках. - М.: Медицина, 1987.
Фомин Н.А., Филин В.П.На пути к спортивному мастерству (адаптация юных спортсменов к физическим нагрузкам). - М.: ФиС, 1986.
Фомин Н.А.Физиология человека: Учеб. пособие для студентов ф-та физ. воспитания пед. ин-тов. - М.: Просвещение, 1982.
Физиология растений
Непосредственно не присоединяется к углероду субстрата дыхания, а идет на биосинтез воды в тканях растения. Если растению давалась вода, меченая по...
Однако прежде всего физиология растений обеспечивает необходимую интеграцию всех биологических...Строение легких ">
Строение легких: а) воздухоносные пути и респираторные отделы; б) легочные альвеолы и их кровоснабжение; в) долька легкого.
Дыхание - совокупность физиологических процессов, обеспечивающих непрерывное поступление кислорода к тканям, использование его в окислительных реакциях, а также удаление из организма образующихся в процессе метаболизма углекислого газа и частично воды. К системе органов дыхания относятся носовая полость , гортань , бронхи и легкие . Дыхание состоит из следующих основных этапов: внешнего дыхания, обеспечивающего газообмен между легкими и внешней средой; газообмена между альвеолярным воздухом и притекающей к легким венозной кровью ; транспорта газов кровью; газообмена между артериальной кровью и тканями; тканевого дыхания.
Вдох и выдох
Легкие через воздухоносные пути сообщаются с атмосферой. При каждом вдохе атмосферный воздух входит в легкие, и при каждом выдохе небольшая часть альвеолярного воздуха выходит в атмосферу. Основная причина газообмена между окружающей средой и альвеолярным воздухом, заполняющим полость легких, - это градиент давления. В момент вдоха давление воздуха в полости легких становится меньше атмосферного и воздух поступает в легкие. При выдохе давление воздуха в легких становится немного выше атмосферного, и воздух из легких выходит в окружающую среду. Изменение давления воздуха в полости легких обусловлено изменением их объема при дыхании. Характеристикой смеси газов, занимающей определенный объем, служит так называемое парциальное давление. Если объем увеличивается - парциальное давление падает, если объем уменьшается - парциальное давление увеличивается.
В легких нет мышечной ткани и нет механизмов, позволяющих легким активно изменять свой объем. Поэтому существует аппарат вентиляции легких, состоящий из грудной клетки (ребра и грудина) и поперечно-полосатых дыхательных мышц. В процессе дыхания аппарат вентиляции вследствие сокращения основных дыхательных мышц совершает ритмические дыхательные движения. При спокойном дыхании в процессе дыхания участвуют поперечно-полосатые наружные межреберные мышцы и диафрагма, которая является главной дыхательной мышцей. При форсированном дыхании в этот процесс могут вовлекаться десятки других поперечно-полосатых мышц туловища.
Три листка плевры.
Легкие окружены париетальной и висцеральной плеврой . Париетальная плевра сращена с грудной клеткой. Висцеральная плевра сращена с легкими. Между париетальной и висцеральной плеврой имеется тонкая щель, заполненная мономолекулярным слоем серозной жидкости. Поверхностное натяжение этой жидкости прочно притягивает друг к другу оба листка плевры, так что при оттягивании одного листка плевры другой следует за ним. Эта ситуация может быть смоделирована, если прижать друг к другу два небольших стекла, капнуть между ними капельку воды и попробовать разъединить. За счет сил поверхностного натяжения воды, их связывающих, разъединить стекла не удастся. Таким образом, во время дыхания грудная клетка тянет за собой париетальную плевру, которая сращена с ней. Париетальная плевра тянет за собой висцеральную плевру, связанную с ней силами поверхностного натяжения серозной жидкости, а висцеральная плевра тянет ткань легких.
Вдох - активный процесс, который совершается именно благодаря сокращению дыхательных мышц. Сокращение наружных межреберных мышц приводит к подъему реберных дуг, грудина отходит немного вперед. Одновременно сокращаются мышечные волокна диафрагмы, ее сухожильный центр смещается книзу, оттесняя брюшные внутренности вперед и вниз. Объем грудной полости увеличивается в трех взаимно перпендикулярных плоскостях. Смещение диафрагмы при спокойном дыхании составляет 1-1,5 см, а при глубоком - может увеличиваться до 10 см. При смещении диафрагмы на 1 см объем грудной полости изменяется примерно на 250-270 мл. Сокращения диафрагмы обуславливают около 50-70% глубины вдоха. Легкие пассивно следуют за всеми изменениями объема грудной полости при сокращении и расслаблении дыхательных мышц. Атмосферное давление, действующее на легкие только со стороны воздухоносных путей, плотно прижимает их к грудной стенке. Атмосферное давление, действующее на нас снаружи, демпфируется кожей и подкожной жировой тканью, поэтому оно не достигает легких с наружной поверхности тела. Грудная полость и плевральная полость, которые окружают легкие, герметичны и с атмосферой не сообщаются.
Одностороннее атмосферное давление, действующее на легкие со стороны дыхательных путей, - главная движущая сила легких. Существуют и другие силы, обусловливающие увеличение объема легких при вдохе и уменьшение при выдохе. Увеличение объема грудной полости в момент вдоха приводит к увеличению объема легких, парциальное давление воздуха в них несколько снижается, и воздух из окружающей среды заходит в легкие.
При выдохе объем грудной клетки уменьшается за счет возврата диафрагмы в исходное состояние и расслабление межреберных мышц. Это приводит к увеличению давления внутри легких, которое превышает атмосферное. Так, попытка сделать сильный выдох, если воздухоносные пути закрыты, вызывает значительный рост давления в альвеолах. В нормальных условиях в результате плавного уменьшения легочного объема создается градиент давления, и воздух пассивно выходит из легких.
В целом, внутренняя поверхность легких связана с атмосферой, а внешняя поверхность легких за счет герметичности грудной клетки такой связи не имеет. Именно это явление позволяет осуществлять вдох и выдох. При нарушении герметичности грудной клетки (например, при ранениях) атмосферное давление начинает действовать не только на внутреннюю поверхность легких, но и на внешнюю. Это приводит к тому, что легкие спадаются (пневмотрокс) и акты вдоха и выдоха становятся невозможными. Двусторонний пневмотрокс, затрагивающий оба легких, если не применяется искусственное нагнетание воздуха в легкие, ведет к смерти.
Газообмен между внешней средой и организмом (три этапа дыхания).
Механизм дыхательных движений (изменение объема грудной клетки) за счет диафрагмы и мышц брюшного пресса (а) и сокращения наружных межреберных мышц (б).
Дыхание состоит из двух фаз: вдох и выдох . Во время вдоха сокращаются мышцы диафрагмы и межреберные мышцы. Диафрагма прогибается вниз, надавливая на органы брюшной полости и увеличивая объем грудной клетки; в результате сокращения межреберных мышц раздвигаются ребра, способствуя еще большему расширению грудной полости. В обычных условиях давление внутри альвеол во время вдоха становиться чуть меньше, чем атмосферное, примерно на -3 мм ртутного столба. Эта разница в давлении заставляет воздух поступать внутрь дыхательных путей, и таким образом уравновешивается. При выдохе происходит обратный процесс: давление в альвеолах увеличивается относительно атмосферного давления на +3 мм ртутного столба, что приводит к выходу содержащегося в них воздуха наружу.
Если на поверхности подводник выполняет произвольную гипервентиляцию, во время усиленного выдоха давление внутри альвеол может увеличиться до +100 мм ртутного столба, а при вдохе оно может упасть до –80 мм ртутного столба. Эти числа объясняют нам, почему вход и выход воздуха из легких напрямую зависит от соотношения показателей атмосферного давления и давления внутри альвеол. Так, например, когда атмосферное давление превышает альвеолярное, воздух будет переходить из области с большим давлением (внешняя) в область с меньшим давлением (альвеолы); и наоборот, если альвеолярное давление больше атмосферного, воздух будет выталкиваться из альвеол наружу.
Рефлекторная регуляция дыхания
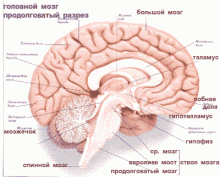 Автоматизм дыхания регулируется клетками центральной нервной системы, объединенными в группы; это так называемые дыхательные центры
, расположенные в определенных зонах мозга: в бульбарной части
продолговатого мозга и варолиевом мосту
.
Автоматизм дыхания регулируется клетками центральной нервной системы, объединенными в группы; это так называемые дыхательные центры
, расположенные в определенных зонах мозга: в бульбарной части
продолговатого мозга и варолиевом мосту
.Бульбарный дыхательный центр разделен на два отдела: инспираторный центр и экспираторный центр, отвечающие соответственно за вдох и выдох; оба они воспринимают химические импульсы, связанные с концентрацией в крови углекислого газа (CO2).
В варолиевом мосту находятся два других дыхательных центра: апнеустический центр и пневмотаксический центр , которые выполняют свои определенные функции в рефлекторной регуляции дыхания и имеют основополагающее значение для погружений на задержке дыхания. Действительно, апнеустический центр – это отправная точка инспираторного импульса, тогда как пневмотаксический центр – это орган, в котором берут свое начало тормозящие импульсы бульбарного инспираторного центра.
Мозговые дыхательные центры: бульбарный дыхательный центр устанавливает дыхательный ритм и чувствительно реагирует на любое увеличение количества CO2, отвечая ускорением дыхания до 6-7 раз.
Химическая регуляция дыхания
Основная задача дыхательной системы – поддерживать в норме уровни содержания CO2 и O2 в крови для обеспечения жизненных функций. Изменение парциального давления этих газов в организме непосредственно влияет на частоту и глубину дыхания.
Из двух этих газов наибольшее влияние на процесс дыхания, несомненно, имеет CO2, поскольку изменение именно его концентрации в крови вызывает реакцию дыхательных центров. Действительно, при любом изменении концентрации CO2 в крови, будь то увеличение или уменьшение, происходит стимуляция бульбарных химических рецепторов , приводящая к раздражению одного из двух центров в варолиевом мосту (апнеустического или пневмотаксического), которые в свою очередь посылают импульсы в один из бульбарных центров (инспираторный или экспираторный).
Из всего вышесказанного становится ясно, что увеличение концентрации CO2 в крови приводит к стимуляции дыхания (возбуждается апнеустический центр и посылает импульсы бульбарному инспираторному центру, который стимулирует дыхание). Поскольку под водой невозможно сделать вдох, увеличение CO2 в крови вызывает диафрагмальные сокращения , характеризуемые серией напряжений и расслаблений мышц диафрагмы – очевидный сигнал тревоги, обозначающий достижение предела задержки дыхания.
 Уменьшение содержания CO2 в крови, как при гипервентиляции, может отсрочить появление дыхательного стимула, поскольку, как было сказано ранее, в бульбарный инспираторный центр импульс поступает только при определенном повышенном уровне концентрации этого газа, достижение которого при гипервентиляции запаздывает, потому что в начале задержки дыхания содержание CO2 в крови сильно занижено. Следовательно, гипервентиляция задерживает сигнал тревоги, используемый организмом для предупреждения о достижении предела задержки. Опасность заключается в том, что, прежде чем уровень CO2 поднимется достаточно для стимуляции дыхания, уровень кислорода может упасть ниже критического уровня. По этой причине гипервентиляция категорически запрещается
; коротко говоря, она значительно понижает в организме уровень защиты и возможность предупреждения об опасности.
Уменьшение содержания CO2 в крови, как при гипервентиляции, может отсрочить появление дыхательного стимула, поскольку, как было сказано ранее, в бульбарный инспираторный центр импульс поступает только при определенном повышенном уровне концентрации этого газа, достижение которого при гипервентиляции запаздывает, потому что в начале задержки дыхания содержание CO2 в крови сильно занижено. Следовательно, гипервентиляция задерживает сигнал тревоги, используемый организмом для предупреждения о достижении предела задержки. Опасность заключается в том, что, прежде чем уровень CO2 поднимется достаточно для стимуляции дыхания, уровень кислорода может упасть ниже критического уровня. По этой причине гипервентиляция категорически запрещается
; коротко говоря, она значительно понижает в организме уровень защиты и возможность предупреждения об опасности.У ныряльщика на задержке дыхания сигналом к подъему является ощущение «кислородного голодания» – дисапноэ , появляющееся вследствие повышения уровня CO2 в крови, которое приводит к раздражению бульбарных химических рецепторов (это особые рецепторы, чувствительные к химическим изменениям крови) с целью стимуляции дыхательного центра для нового вдоха. Чтобы продлить задержку дыхания, подводник иногда сдерживает первые признаки дисапноэ, но это может привести к опасным последствиям, особенно, если подводник, как это обычно бывает, гипервентилирует легкие, ошибочно полагая, что увеличивает таким образом свой запас кислорода.
На самом деле гипервентиляция приводит к уменьшению CO2 в альвеолах и крови, что, как мы впоследствии увидим, повышает риск гипоксии (чрезмерное уменьшение парциального давления PpO2) и вызывает у человека гипоксический обморок .
В нормальных условиях перепады парциального давления O2 и CO2 в крови и в альвеолярном воздухе способствуют прохождению O2 из легких в кровь, и CO2 из крови в легкие. Во время погружения увеличение давления внутри легких способствует распространению O2, но и препятствует выходу CO2. Действительно, на глубине 10 метров внутрилегочное давление таково, что CO2 перемещается в обратном направлении: из легких в кровь, а не из крови в легкие. На глубине запас O2 в легких уменьшается гораздо быстрее, чем на поверхности, и одновременно повышается PCO2. Таким образом, сигнал к всплытию появится с запозданием относительно реального остатка кислорода, а это может вызвать у неопытного подводника, плохо знающего собственные возможности, иллюзию, что можно и дальше задерживать дыхание.
Во время всплытия давление газа быстро падает, как в легких, так и в крови. При уменьшении давления O2 до гипоксичного уровня у подводника может произойти потеря сознания с последующим обмороком и возникновением риска утопления.
Опасность еще больше увеличивается, если на поверхности подводник делал гипервентиляцию, поскольку, как мы уже видели, эта методика дает лишь небольшое увеличение парциального давления кислорода, а по большей части происходит значительное понижение парциального давления углекислого газа. Это приводит к последующему запаздыванию стимуляции дыхательных центров, дающих сигнал тревоги о приближении предела задержки дыхания, который позволяет вовремя вернуться на поверхность для дыхания.
По материалам книги Марко Барди "Учебник подводной охоты на задержке дыхания"











