Funzioni di diverse parti del cervello. La struttura del cervello umano. Struttura della corteccia cerebrale
Cervello, encefalo, è posto nella cavità cranica ed ha una forma a schema generale corrispondente ai contorni interni della cavità cranica. La sua superficie superolaterale, o dorsale, secondo la volta cranica, è convessa, e il lato ventrale, o base del cervello, è più o meno appiattito e irregolare. Il cervello può essere diviso in tre grandi parti: gli emisferi grande cervello(hemispheriae cerebri), cervelletto (cervelletto) e tronco encefalico (truncus cerebri). Maggior parte L'intero cervello è occupato dagli emisferi, seguiti per dimensioni dal cervelletto, il resto, una parte relativamente piccola, è il tronco encefalico.
Vero sistemi complessi possono essere modellati matematicamente come grafici che rivelano le proprietà del sistema. Proponiamo due nuovi diagrammi cerebrali, uno di 264 presunte regioni funzionali e l'altro una modifica delle reti voxel che elimina le relazioni potenzialmente artificiali a breve distanza. Questi grafici contengono molti sottografi che sono in buon accordo con i funzionali noti sistemi cerebrali. Altri sottografi mancano di identità funzionali stabilite; proponiamo possibili caratteristiche funzionali per questi sottografi.
cervello. Entrambi gli emisferi sono separati l'uno dall'altro da una fessura, fisstira longitudinalis cerebri, andando in direzione sagittale. Nelle profondità della fessura longitudinale, gli emisferi sono collegati tra loro da aderenze - corpo calloso, corpo calloso e altre formazioni sottostanti. Davanti al corpo calloso passa la fessura longitudinale, e dietro passa nella fessura trasversale del cervello, fissiira trasversale cerebrale, separando le parti posteriori degli emisferi dal cervelletto sottostante.
Inoltre, le misurazioni del grafico della rete areale mostrano che il sottografo della modalità predefinita condivide le proprietà della rete con i sottografi sensoriali e motori: è internamente integrato ma isolato dagli altri sottografi, simile a un sistema di elaborazione. Il grafico voxel modificato rivela anche motivi spaziali nei modelli dei sistemi in tutta la corteccia.
Negli ultimi anni i progressi nel neuroimaging, che facilitano lo studio delle relazioni cerebrali negli esseri umani, hanno suscitato enorme interesse da parte di scienziati e medici. Queste correlazioni sono di grande interesse per la comunità medica perché tutti numero maggiore condizioni patologiche, a quanto pare, si riflette nella connettività funzionale tra le singole regioni del cervello.
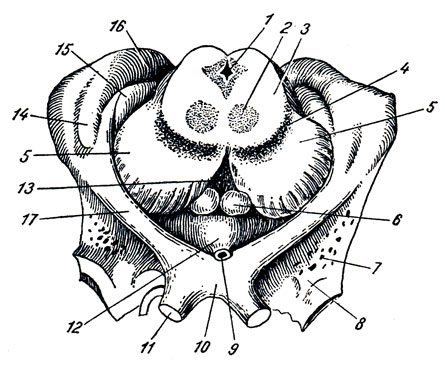
Superficie inferiore del cervello(Fig. 272). Dal lato della superficie inferiore del cervello, il cervello inferiore sfuma, non è visibile solo il lato inferiore degli emisferi cerebrali e del cervelletto, ma anche l'intera superficie inferiore del tronco encefalico, nonché i nervi che si estendono dal cervello. Qui incontriamo le seguenti parti, se andiamo dalla parte anteriore a quella posteriore. La sezione anteriore della superficie inferiore del cervello è rappresentata dai lobi frontali degli emisferi. Sulla superficie inferiore dei lobi frontali si notano bulbi olfattivi, btilbi olfattivi, ai quali sottili filamenti nervosi, fila olfattiva, che insieme formano il primo paio di nervi della testa, si avvicinano dalla cavità nasale attraverso le aperture della lamina cribrosa dell'osso etmoidale - nervi olfattivi, nn. olfattivi. Di solito, quando si rimuove il cervello dal cranio, questi fili vengono strappati dal billbus olfactorius. I bulbi olfattivi continuano posteriormente nell'interno tratti olfattivi , tratto olfattivo, ciascuna terminante con due radici, tra le quali c'è un'elevazione chiamata trigono olfattivo. Direttamente dietro quest'ultimo su entrambi i lati c'è sostanza perforata anteriore, sostanza perforata anteriore, così chiamato a causa dei piccoli fori che si trovano qui attraverso i quali i vasi sanguigni passano nel midollo.
Mielencefalo, midollo allungato
In questa struttura un sistema complesso formalizzato come un oggetto matematico costituito da un insieme di elementi e un insieme di relazioni a coppie tra elementi. Gli elementi sono chiamati nodi, le relazioni sono chiamate collegamenti e gli insiemi di questi nodi con le loro connessioni sono chiamati grafici o reti. Un elenco breve e parziale di argomenti consolidati nella teoria dei grafi include la quantificazione della gerarchia e della sottostruttura in un grafo, l'identificazione di hub e nodi critici, la determinazione della facilità con cui il traffico scorre in parti differenti e a diverse scale di rete e valutazione della controllabilità del sistema.
Nel mezzo tra entrambi gli spazi perforati anteriori si trova chiasma ottico, chiasma ottico a forma di lettera "X". Una lamina sottile si estende dalla superficie superiore del chiasma grigio, lamina terminale, andando più in profondità nella fissura longitudinalis cerebri. Posto dietro il chiasma ottico protuberanza grigia, tubero cinereo; la sua sommità è allungata in un tubo stretto, il cosiddetto imbuto, infundibolo, sul quale, come una bacca su un gambo, pende un corpo tondeggiante - un'appendice del cervello, ipofisi cerebrale. Quando il cervello è posizionato al suo posto nel cranio, l'appendice viene posizionata nella cavità della sella turcica. Dietro il tumulo grigio ce ne sono due sferici bianco elevazioni - corpi mastoidei, corpi mamillari. Dietro di loro si trova una fossa quadrangolare piuttosto profonda, fossa interpeduncolare, delimitata lateralmente da due fitti cespi bianchi convergenti posteriormente e chiamati gambe del cervello, peduncoli cerebrali. Il fondo della fossa è forato con fori per i vasi, e per questo si chiama sostanza perforata posteriore, sostanza perforata posteriore. Accanto a questa sostanza, nel solco del bordo mediale del peduncolo cerebrale, fuoriesce su entrambi i lati III coppia - nervo oculomotore, N. oculomotorio. Il più sottile dei nervi cerebrali è visibile sul lato dei peduncoli cerebrali nervo trocleare , N. trocleare- IV paio, che però non nasce dalla base del cervello, ma dalla sua parte dorsale, dal cosiddetto velo midollare superiore. Dietro i peduncoli cerebrali c'è uno spesso albero trasversale: il ponte, ponte(Varolii), che, rastremandosi lateralmente, si immerge nel cervelletto. Vengono chiamate le parti laterali del ponte più vicine al cervelletto peduncolo cerebellare medio, peduncoli cerebellari medii; al confine tra loro e il ponte stesso emerge su entrambi i lati una coppia di V - nervo trigemino, N. trigemino. Dietro il ponte si trova midollo, midollo allungato; tra esso e il bordo posteriore del ponte ai lati della linea mediana è visibile l'inizio della VI coppia - nervo abducente, N. abducente; ancora più lateralmente, in corrispondenza del bordo posteriore dei peduncoli cerebellari medi, emergono affiancati su entrambi i lati altri due nervi: VII paio - nervo facciale, N. facialis, e VIII coppia - n. vestibolococleare.
Ostacolo attuale basato sullo studio grafico organizzazione funzionale del cervello è che è molto difficile identificare i singoli nodi che compongono la rete cerebrale. Secondo i primi principi del trattamento di un grafico come modello di un sistema reale, se i nodi del grafico non rappresentano accuratamente gli elementi reali del sistema, il grafico stesso è un modello distorto e le proprietà teoriche del grafico differiranno da quelle reali. le vere proprietà del sistema. Il cervello è una rete complessa con organizzazione macroscopica a livello di aree funzionali e nuclei sottocorticali, ma il numero e la localizzazione di queste entità è in gran parte sconosciuto nell’uomo.
Tra la piramide e l'olivo midollo allungato le radici stanno uscendo XII coppie - nervo ipoglosso, N. ipoglosso. Coppie di radici IX, X e XI - N. glossofaringeo, n. vago e n. accessorio(parte superiore) - emergono dal solco dietro l'oliva. Le fibre inferiori della coppia XI partono già dal midollo spinale nella sua parte cervicale.
Sviluppo (embriogenesi) del cervello
Il tubo midollare viene diviso molto presto in due sezioni corrispondenti al cervello e al midollo spinale. La sua sezione anteriore ed espansa, che rappresenta, come già accennato, il rudimento del cervello, è sezionata mediante costrizioni in tre vescicole midollari primarie disposti uno dietro l'altro: anteriore, prosencefalo, medio, mesencefalo e posteriore, rombencefalo. La vescicola cerebrale anteriore è chiusa davanti dalla cosiddetta placca terminale, lamina terminalis. Questo stadio di tre vescicole, dopo successiva differenziazione, si trasforma in cinque vescicole, dando origine alle cinque parti principali del cervello (Fig. 273). Allo stesso tempo, il tubo cerebrale si piega in direzione sagittale. Innanzitutto, nella regione della vescicola media, si sviluppa una curva parietale convessa posteriormente, e poi, al confine con il rudimento del midollo spinale, si sviluppa anche una curva occipitale convessa dorsalmente. Tra di loro si forma una terza curva nella regione della vescicola posteriore, convessa verso il lato ventrale (curva pontina).
Lo scopo principale di questo rapporto è quello di almeno, superando parzialmente questo ostacolo. Abbiamo sviluppato metodi per identificare, per quanto possibile, un insieme di nodi più adatti e definire una rete basata su questi nodi. Proponiamo anche una seconda nuova rete cerebrale basata su una modifica degli approcci basati su voxel e consideriamo alcune delle sue proprietà rispetto al primo grafico.
Esistono molti metodi utilizzati per suddividere reti di grandi dimensioni in sottoreti con nodi altamente correlati, in modo che i nodi all'interno dei sottografi siano più densamente connessi tra loro rispetto al resto del grafico. Pertanto, la presenza di sottografi corrispondenti a sistemi funzionali indica che il grafico modella accuratamente alcune caratteristiche dell'organizzazione cerebrale, mentre l'assenza di tali sottografi solleva il sospetto che il grafico possa essere sottospecificato.
Attraverso quest'ultima curva vescicola midollare posteriore, rombencefalo, è diviso in due sezioni. Di questi, quello posteriore, il mielencefalo, si trasforma nel midollo allungato durante lo sviluppo finale, e da sezione anteriore, detto recintofalo, si sviluppa sul lato ventrale del ponte e sul lato dorsale del cervelletto. Il metencefalo è separato dalla vescicola mesencefalica che gli sta di fronte da una stretta costrizione, l'istmo rombencefalico. La cavità comune del rombencefalo, che ha forma di rombo nella sezione frontale, forma il IV ventricolo, comunicante con il canale centrale del midollo spinale. Le sue pareti ventrali e laterali, a causa dello sviluppo in esse dei nuclei dei nervi cefalici, si ispessiscono notevolmente, mentre la parete dorsale rimane sottile. Nella regione del midollo allungato la maggior parteè costituito da un solo strato epiteliale, fuso con la coroide (tela chorioidea inferiore). bolla media, mesencefalo, si addensano con lo sviluppo della materia cerebrale in essi in modo più uniforme. Ventralmente da essi derivano i peduncoli cerebrali e, sul lato dorsale, la placca quadrigeminale (vedi Fig. 273). La cavità della vescicola media si trasforma in uno stretto canale: un acquedotto, che si collega al ventricolo IV.
Sulla base di questa ipotesi, apriamo questo articolo esaminando le strutture dei sottografi di quattro grafici su scala mondiale in un unico set di dati. Come accennato in precedenza, vengono studiati due nuovi grafici: un grafico delle regioni funzionali dedotte e una modifica delle reti di voxel che eliminano le correlazioni a breve distanza. Per fare un confronto, vengono utilizzati altri due grafici standard: un grafico a particelle tratto da un popolare atlante del cervello e un grafico voxel standard. Per prevedere i risultati, i sottografi nella rete areale sono significativamente più simili a sistemi funzionali rispetto ai sottografi in un grafico basato su atlante, e i sottografi in una rete voxel modificata sono più simili ai sistemi funzionali rispetto a una rete voxel standard.
subisce differenziazioni e modifiche formali più significative vescicola midollare anteriore, prosencefalo, che è suddiviso in Indietro, diencefalo (diencefalo) e anteriore, telencefalo ( telencefalo). Le pareti laterali del diencefalo, ispessendosi, formano i tubercoli visivi (talami). Inoltre le pareti laterali, sporgenti ai lati, formano due vescicole ottiche, dalle quali successivamente si sviluppano la retina degli occhi e i nervi ottici. La parete dorsale del diencefalo rimane sottile, sotto forma di placca epiteliale fusa con la coroide (tela chorioidea superior). Nella parte posteriore, da questo muro si forma una sporgenza, a causa della quale si forma l'epifisi (corpo pineale). Le gambe cave delle vescicole ottiche vengono retratte dal lato ventrale nella parete della vescicola cerebrale anteriore, a seguito della quale si forma una depressione sul fondo della cavità di quest'ultima, recessus opticus, la cui parete anteriore è costituita di una sottile lamina terminale. Dietro il recesso ottico appare un'altra depressione a forma di imbuto, le cui pareti danno origine al tuber cinereum, all'infundibolo e al lobo posteriore (nervoso) dell'ipofisi cerebrale. Ancora più indietro, nella zona del diencefalo, si formano corpi mamillari accoppiati sotto forma di un'unica elevazione. Si forma la cavità del diencefalo III ventricolo.
Confronto di reti: identificazione di quattro reti cerebrali
Dati questi risultati, conduciamo tutta la linea ulteriori analisi su nuovi grafici per conoscere meglio l'organizzazione funzionale del cervello, con alcuni nuovi ed interessanti risultati. Due nuovi e due metodi standard le definizioni dei grafici sono state esaminate in un'ampia coorte di giovani adulti sani. Per ribadire, i grafici sono costituiti da un insieme di nodi e da un insieme di connessioni tra i nodi. Pertanto, la matrice di correlazione incrociata di un insieme di nodi definisce un grafico. Poiché non esiste una soglia “corretta”, tutte le analisi sono state eseguite su un intervallo di valori di soglia, in genere a partire da circa il 10% della densità connettiva e aumentando fino a quando le reti diventavano altamente frammentate.
Il telencefalo è diviso in una parte media, più piccola (telencefalo medio) e due grandi parti laterali: le vescicole degli emisferi cerebrali (hemispherium dextrum et sinistrum), che nell'uomo crescono molto fortemente e alla fine dello sviluppo le loro dimensioni superano significativamente la resto del cervello. La cavità del mezzo telencefalico, che è continuazione anteriore della cavità del diencefalo (III ventricolo), comunica lateralmente attraverso i fori interventricolari con le cavità delle vescicole degli emisferi, che si trovano sul cervello sviluppato sono chiamati ventricoli laterali. La parete anteriore della parte media del telencefalo medio, che è diretta continuazione della lamina terminale, all'inizio del primo mese di vita embrionale forma un ispessimento, la cosiddetta placca commissurale, da cui si corpo calloso e commissura anteriore.
Il primo nuovo programma è stato definito secondo principi neurobiologici. Il cervello è una rete complessa con un'organizzazione spaziale e funzionale gerarchica a livello di neuroni, circuiti locali, colonne, aree funzionali e sistemi funzionali. I centri delle presunte regioni sono stati determinati utilizzando due metodi indipendenti che operano su set di dati che non sono stati utilizzati nell'analisi del grafico. La combinazione di questi metodi ha prodotto 264 presunte regioni che abbracciano la corteccia cerebrale, le strutture sottocorticali e il cervelletto.
Alla base delle vescicole degli emisferi si forma all'interno su entrambi i lati una sporgenza, la cosiddetta tubercolo nodulare, da cui si sviluppa lo striato, corpo striato. Parte della parete mediale della vescicola degli emisferi rimane sotto forma di un unico strato epiteliale, che viene arrotolato nella vescicola da una piega della coroide (plesso corioideo). Sul lato inferiore di ciascuna vescicola dell'emisfero, già nella quinta settimana di vita embrionale, si forma una sporgenza: il rudimento cervello olfattivo, rinencefalo, che è gradualmente delimitato dalla parete degli emisferi da un solco corrispondente alla fessura rinalis lateralis. Con lo sviluppo della materia grigia (corteccia), e poi della materia bianca nelle pareti dell'emisfero, quest'ultima aumenta e forma il cosiddetto mantello, pallio, che giace sopra il cervello olfattivo e copre non solo le collinette visive, ma anche il superficie dorsale del mesencefalo e del cervelletto.
Le regioni di interesse sono state modellate come sfere con un diametro di 10 mm. Questa procedura ha prodotto grafici di presunte aree funzionali in cui ciascun nodo rappresentava, per quanto possibile, un elemento dell'organizzazione cerebrale. Il secondo nuovo grafico considerato è stata una modifica delle reti voxel in cui sono state eliminate tutte le connessioni a breve distanza. Questa modifica è nata da diverse osservazioni pratiche. Innanzitutto, i voxel vicini condividono il segnale non biologico come risultato di inevitabili passaggi nell’elaborazione dei dati.
In terzo luogo, come si vedrà tra breve, i grafici dei voxel sono dominati a soglie più elevate da relazioni a breve distanza che sono logicamente parzialmente artificiali in base alle considerazioni di cui sopra. Vengono presentate reti di canali voxel modificate in cui sono esclusi tutti i collegamenti che terminano entro 20 mm dal nodo di origine, sebbene siano state testate anche altre distanze con risultati simili.
Man mano che l'emisfero cresce, aumenta prima nel lobo frontale, poi nei lobi parietale e occipitale e infine nel lobo temporale. Ciò fa sembrare che il mantello ruoti attorno alle cuspidi visive, prima dalla parte anteriore a quella posteriore, poi verso il basso e infine curvandosi in avanti verso il lobo frontale. Di conseguenza, sulla superficie laterale dell'emisfero, tra il lobo frontale e il lobo temporale che si avvicina ad esso, si forma un foro, fossa cerebri lateralis (Sylvii), che, quando i lobi nominati del cervello sono completamente riuniti, gira in uno spazio vuoto, sulcus cerebri lateralis (Sylvii); al suo fondo si forma un'isola, insula.
Reti a confronto: corrispondenza tra sottografi e sistemi funzionali
Due metodi standard di generazione di grafici sono basati su batch e basati su voxel. Questo atlante divide la corteccia e le strutture sottocorticali in regioni basate su punti di riferimento anatomici. Questo algoritmo utilizza l'equazione della mappa per ridurre al minimo le descrizioni teoriche delle passeggiate casuali in un grafico.
Struttura del cervello
Abbiamo previsto che i grafici ben formati avrebbero sottografi ben formati corrispondenti ai principali sistemi cerebrali funzionali. La Figura 3 fornisce una panoramica del modo in cui ciascuna rete ha soddisfatto questa previsione. Le tre colonne successive mostrano i sottografi di una soglia di analisi per ciascun grafico. La seconda colonna mostra insieme sia le assegnazioni fisse che quelle dei voxel poiché sono in ottimo accordo. I grafici di zona e di voxel modificato contengono sottografi corrispondenti a ciascuno dei sistemi funzionali e questi sottografi contengono la maggior parte o tutte le regioni cerebrali coinvolte nei sistemi funzionali e talvolta alcune regioni cerebrali aggiuntive.
Durante lo sviluppo e la crescita dell'emisfero, insieme ad esso, la "rotazione" indicata e le sue camere interne, i ventricoli laterali del cervello (resti della cavità della vescica primaria), nonché parte del corpo striato (caudato nucleo) sviluppano e compiono la “rotazione” indicata, il che spiega la somiglianza della loro forma con la forma dell'emisfero: i ventricoli hanno parti anteriore, centrale e posteriore e una parte inferiore che curva verso il basso e in avanti (vedi Fig. 295). ; il nucleo caudato ha una testa, un corpo e una coda che curva verso il basso e in avanti.
Selezione delle definizioni di rete per ulteriori analisi
Il grafico voxel standard descrive bene alcuni sistemi funzionali, ma altri non sono completamente rappresentati. I grafici di zona e di voxel modificati corrispondono meglio alle nostre previsioni sulla corrispondenza tra sistemi funzionali e sottografi nelle reti cerebrali. Passiamo ora dalla nostra attenzione sui risultati confermativi a nuove osservazioni sull'organizzazione funzionale del cervello che possono essere tratte da grafici voxel areali e modificati. Continueremo a concentrarci sulla rete a livello di sottografo.
Solchi e circonvoluzioni(Fig. 274, 275, 276) sorgono a causa della crescita irregolare del cervello stesso (D. Zernov), che è associata allo sviluppo delle sue singole parti. Così, al posto del cervello olfattivo, sorgono il sulcus olfactorius, il sulcus hyppocàmpi e il sulcus cinguli; al confine delle estremità corticali della pelle e analizzatore motorio (il concetto di analizzatore e la descrizione dei solchi, vedi sotto) - sulcus centralis; al confine tra l'analizzatore motorio e la zona premotoria, che riceve impulsi dai visceri - sulcus precentralis; sul posto analizzatore uditivo- solco temporale superiore; nell'area dell'analizzatore visivo - sulcus calcarinus e sulcus parieiooccipitdlis.
Inizieremo con una discussione sulle identità dei sottografi, poi esamineremo le relazioni e le proprietà di sottografi specifici e termineremo con osservazioni sulle relazioni tra tutti i sottografi. Le identità dei sottografi rosso, giallo, verde e tigre sono già chiare. Vengono ora considerati i rimanenti sottografi principali.
Diversi sottografi corrispondono alle regioni sensoriali e motorie. A soglie moderate, la corteccia somatosensoriale-motoria era divisa in sottografi dorsale e ventrale. Tre dati indicano le identità di questi sottografi. Innanzitutto, le coordinate riportate per la rete di controllo del compito sono coordinate di salienza dorsale isolate, sebbene la maggior parte delle altre coordinate non discriminino tra sistemi funzionali concorrenti. Ipotizziamo che il sottografo viola rappresenti più da vicino un sistema di controllo del compito violoncello-opercolare, mentre il sottografo nero rappresenti più da vicino un sistema di salienza, sebbene le prove di tali assegnazioni siano preliminari.
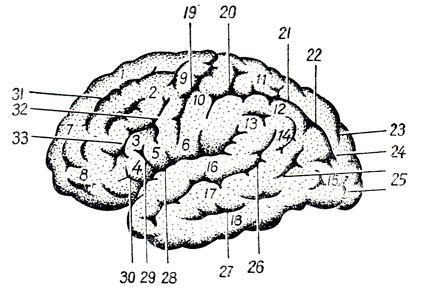
Tutti questi solchi, che compaiono prima degli altri e sono caratterizzati da assoluta costanza, appartengono, secondo D. Zernov, a solchi della prima categoria. I restanti solchi, che hanno nomi e nascono anch'essi in relazione allo sviluppo degli analizzatori, ma compaiono un po' più tardi e sono meno costanti, appartengono a solchi della seconda categoria. Al momento della nascita sono presenti tutti i solchi della prima e della seconda categoria. Infine, numerosi piccoli solchi che non hanno nome compaiono non solo nella vita uterina, ma anche dopo la nascita. Sono estremamente variabili nel tempo di apparizione, nel luogo e nel numero; Questo solchi di terza categoria. La diversità e la complessità del rilievo cerebrale dipende dal grado del loro sviluppo. Altezza cervello umano V periodo embrionale e nei primi anni di vita, mentre il corpo cresce rapidamente, adattandosi a un nuovo ambiente, acquisendo la capacità di camminare eretto e la formazione di un secondo, verbale, sistema di segnalazione, si manifesta in modo molto intenso e termina all'età di 20 anni. Nei neonati, il cervello (in media) pesa 340 g nei ragazzi e 330 g nelle ragazze, e in un adulto pesa 1375 g negli uomini e 1245 g nelle donne.
Sono stati trovati anche almeno tre sottografi distribuiti con identificatori funzionali precedentemente sconosciuti. Il primo sottografo comprende parti della mascella posteriore, della corteccia parietale mediale posteriore e parietale laterale. Un altro sottografo distribuito è stato trovato nel frontale, parietale e corteccia temporale a soglie più elevate dell'analisi voxel modificata.
Un altro nuovo sottografo è mostrato in prugna con rappresentazione nella corteccia fusiforme, nella corteccia parietale posteriore preliminale, laterale e mediale e nella corteccia frontale superiore. Passiamo ora dallo studio dei singoli sottografi agli insiemi di sottografi e alle loro relazioni reciproche.
Parti del cervello
In base allo sviluppo embrionale, come già indicato, il cervello è suddiviso in sezioni situate, a partire dall'estremità caudale, nel seguente ordine:
1) rombencefalo, romboide o rombencefalo, che, a sua volta, è costituito da: a) mielencefalo, midollo allungato eb) metencefalo - il rombencefalo stesso; 2) mesencefalo - mesencefalo; 3) prosencefalo, il proencefalo, in cui si distinguono: a) diencefalo - diencefalo eb) telencefalo - telencefalo.
Il “sistema compito-positivo” è costituito da diversi sottografi e il “sistema compito-negativo” è costituito da un sottografo. Questi risultati hanno diverse implicazioni. Dal punto di vista della teoria dei grafi, i sistemi sensoriali e motori e il sistema della modalità predefinita hanno livelli simili di autointegrazione e autocondensazione. Dalla prospettiva del controllo cognitivo sopra descritta, queste somiglianze suggeriscono che il sistema di default opera più come un “sistema di elaborazione” che come un “sistema di controllo”.
I sistemi funzionali sono organizzati secondo schemi topologici in tutta la corteccia
La Figura 7 mostra la presenza di motivi ad una soglia dell'analisi del voxel modificata. Per ciascun sottografo viene costruita la distribuzione delle sue interfacce spaziali con altri sottografi e questi sottografi vicini vengono quindi esaminati per garantire che sia improbabile che essi stessi interagiscano. Ad esempio, il sottografo azzurro interagisce prevalentemente con i sottografi rosso e giallo, che sono essi stessi piccole parti dei confini l'uno dell'altro, implicando un motivo blu-rosso del giallo.
Oltre a queste sezioni, esiste anche un istmo, l'istmo rombencefalo, tra il rombencefalo e il mesencefalo.
Il mesencefalo e il prosencefalo costituiscono il cervello grande, il cervello, in contrasto con il cervello piccolo, il cervelletto - cervelletto.
Rombencefalo
Mielencefalo, midollo allungato
Midollo allungato, cervello, midollo allungato(Fig. 277, 278), rappresenta una continuazione diretta del midollo spinale nel tronco encefalico e fa parte del romboide o romboencefalo. Combina le caratteristiche strutturali del midollo spinale e dipartimento primario testa, che giustifica il suo nome: mielencefalo. Il midollo allungato ha l'aspetto di un bulbo, bulbus cerebri, s. bulbus medullae spinalis (da qui il termine “disturbi tabloid”); l'estremità superiore espansa confina con il ponte e il suo bordo inferiore è il punto di uscita delle radici della prima coppia nervi cervicali o il livello del forame magno.
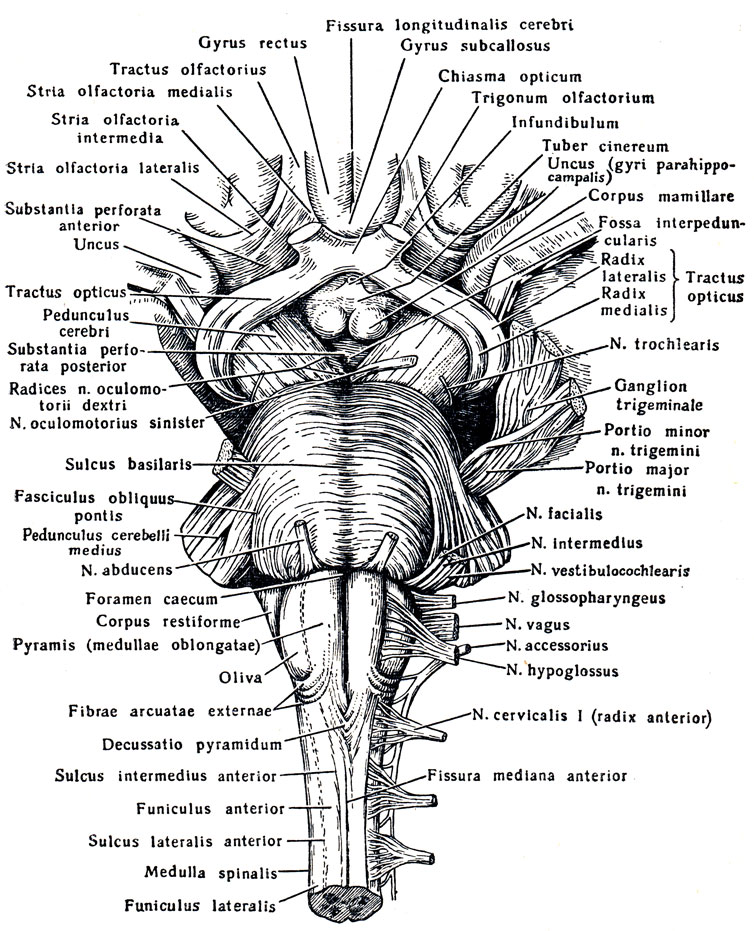
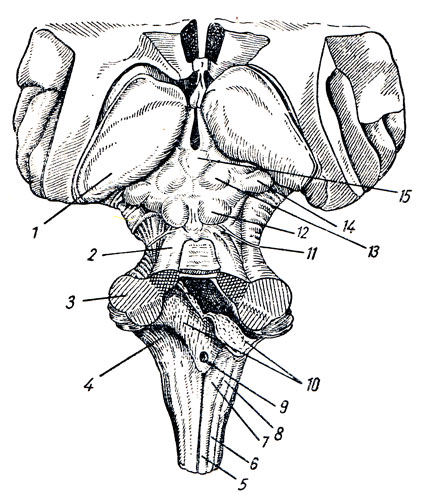
1. Sulla superficie anteriore (ventrale). La fessura mediana anteriore corre lungo la linea mediana del midollo allungato, formando una continuazione dell'omonimo solco del midollo spinale. Ai suoi lati, su entrambi i lati, ci sono due fili longitudinali: piramidi, midollo allungato della piramide, che sembrano continuare nelle corde anteriori del midollo spinale. I fasci di fibre nervose che compongono la piramide si intersecano parzialmente nella profondità della fessura mediana anteriore con fibre simili del lato opposto - decussidtio Pyramidum, dopo di che scendono nella corda laterale sull'altro lato del midollo spinale - tractus corticospinal (pyramidalis) lateralis, in parte rimangono non incrociati e scendono nella corda anteriore del midollo spinale sul suo lato - tratto corticospinale (pyramidalis) anteriore.
Le piramidi sono assenti nei vertebrati inferiori e compaiono durante lo sviluppo della neocorteccia; pertanto, sono più sviluppati nell'uomo, poiché le fibre piramidali collegano la corteccia cerebrale, raggiungendo l'uomo massimo sviluppo, con i nuclei dei nervi cerebrali e le corna anteriori del midollo spinale.
Lateralmente alla piramide si trova un'elevazione ovale - oliva, olfva, che è separata dalla piramide da un solco, sulcus lateralis anterior.
2. Sul lato dorsale (posteriore). Il midollo allungato (vedi Fig. 278) si estende sul solco medio posteriore, una continuazione diretta dell'omonimo solco del midollo spinale. Sui suoi lati si trovano le corde posteriori, limitate lateralmente su entrambi i lati dal solco laterale posteriore debolmente definito. Nella direzione verso l'alto, le corde posteriori divergono lateralmente e vanno al cervelletto, diventando parte della parte inferiore delle sue gambe, pedunculi cerebellares inferiores, confinanti con la fossa romboidale sottostante. Ciascun cordone posteriore è diviso dal solco intermedio in mediale, fascicolo gracile, e laterale, fascicolo cuneato. Nell'angolo inferiore della fossa romboidale, le corde tenere e a forma di cuneo acquisiscono ispessimenti: tuberculum nuclei gracilis e tuberculum nuclei cunedti. Questi ispessimenti sono causati dai nuclei della materia grigia, il nucleo gracile e il nucleo cuneatus, che sono simili alle corde. In questi nuclei terminano le fibre ascendenti delle radici dorsali del midollo spinale (fasci di Gaulle e Burdach) che passano nelle corde posteriori. La superficie laterale del midollo allungato, situata tra il solco lateralis posteriore e anteriore, corrisponde al cordone laterale. Le coppie XI, X e IX di nervi della testa emergono dal solco laterale posteriore dietro l'olivo. Il midollo allungato contiene Parte inferiore fossa romboidale (Fig. 279; vedere Fig. 283).
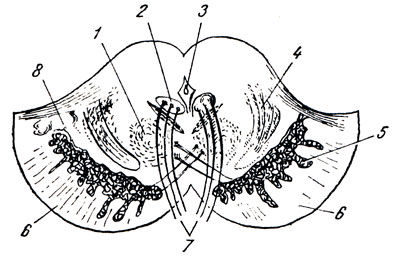
Struttura interna del midollo allungato. Il midollo allungato è nato in connessione con lo sviluppo degli organi statici e acustici, nonché in connessione con l'apparato branchiale legato alla respirazione e alla circolazione sanguigna. Pertanto contiene nuclei della materia grigia, legato all'equilibrio e alla coordinazione dei movimenti, nonché alla regolazione del metabolismo (Fig. 280).
1. Nucleo olivdris, nocciolo di oliva, ha l'aspetto di una lamina contorta di sostanza grigia, aperta medialmente (ilo), e provoca la sporgenza dell'olivo dall'esterno. È associato al nucleo dentato del cervelletto ed è il nucleo intermedio dell'equilibrio, più pronunciato nell'uomo, posizione verticale che richiede l'apparato di equilibrio più perfetto. (C'è anche l'accesso nucl. olivaris, med.)
2. Formatio reticularis, formazione reticolare, formato dall'intreccio delle fibre nervose e di quelle che si trovano tra loro cellule nervose.
3. Nuclei di quattro paia di nervi cefalici inferiori (XII-IX), legati all'innervazione dei derivati dell'apparato branchiale e dei visceri.
4. Centri vitali di respirazione e circolazione associati ai nuclei del nervo vago. Pertanto, se il midollo allungato è danneggiato, può verificarsi la morte.
materia bianca Il midollo allungato contiene fibre lunghe e corte. Quelli lunghi comprendono i tratti piramidali discendenti che attraversano le corde anteriori del midollo spinale, intersecandosi in parte nella zona delle piramidi. Inoltre, dai nuclei dei funicoli posteriori (nucleus gracilis et cuneatus) partono i secondi neuroni delle vie sensoriali ascendenti, che vanno dal midollo allungato al talamo visivo, trdctus bulboihaldmicus. Le fibre di questo fascio formano un'ansa mediale, lemniscus medlis, che si incrocia nel midollo allungato, decussedtio lemniscorum, e sotto forma di un fascio di fibre situato dorsalmente alle piramidi, tra le olive - lo strato dell'ansa interolive - va oltre. Pertanto, nel midollo allungato ci sono due incroci di lunghe vie: quella motoria ventrale, decussatio Pyramidum, e quella sensoriale dorsale, decussatio lemniscorum.
A scorciatoie Questi includono fasci di fibre nervose che collegano i singoli nuclei della materia grigia, nonché i nuclei del midollo allungato con le parti vicine del cervello. Tra questi sono da segnalare il trdctus olivocerebellars ed il fasciculus longitudindlis medlis dorsali allo strato interolivo.
I rapporti topografici delle principali formazioni del midollo allungato sono visibili in una sezione trasversale presa a livello degli olivi (vedi Fig. 280). Le radici che si estendono dai nuclei dei nervi ipoglosso e vago dividono il midollo allungato su entrambi i lati in tre regioni: posteriore, laterale e anteriore. Nella parte posteriore ci sono i nuclei del funicolo posteriore e dei peduncoli cerebellari inferiori, nella parte laterale ci sono il nucleo dell'olivo e la formatio reticularis, e nella parte anteriore ci sono le piramidi.
Metencefalo
Il metencefalo è costituito da due parti: quella ventrale - il ponte e quella dorsale - il cervelletto.
Ponte
Ponte, ponte(Varolii), è uno spesso fusto bianco che si estende lateralmente alla base del cervello, confinante caudalmente con l'estremità superiore del midollo allungato e cranialmente con i peduncoli cerebrali (vedere Fig. 277). Il bordo laterale del ponte è una linea tracciata artificialmente attraverso le radici dei nervi trigemino e facciale, linea trigeminofacidlis. Lateralmente a questa linea si trovano i peduncoli cerebellari medi, peduncoli cerebellari medii, immergendosi su entrambi i lati nel cervelletto. La superficie dorsale del ponte non è visibile dall'esterno, poiché è nascosta sotto il cervelletto, formando parte in alto fossa romboidale (fondo del quarto ventricolo). La superficie ventrale del ponte è di natura fibrosa e le fibre generalmente corrono trasversalmente e sono dirette nei peduncoli cerebellari medii. Lungo la linea mediana della superficie ventrale vi è un dolce solco, sulcus basilaris, in cui si trova a. basilaris.
Struttura interna del ponte. Sulle sezioni trasversali del ponte puoi vedere che è costituito da un ponte inferiore più grande, o parte ventrale, pars ventralis pontis e più piccolo dorsale, pars dorsale pontis. Il confine tra loro è uno spesso strato di fibre trasversali: il corpo trapezoidale, corpo trapezoidale, le cui fibre appartengono a percorso uditivo. Nella zona del corpo trapezoidale si trova un nucleo che è anche correlato alla via uditiva: nucleo dorsalis corporis trapezoidei o nucleo olivaris superiore (BNA). Quest'ultimo nome è stato dato al nucleo perché ha una forma frastagliata, simile al nucleo del midollo allungato olivare.
Pars ventralis contiene fibre longitudinali e trasversali, tra le quali sono sparsi i nuclei propri della materia grigia, nuclei pontis.
Le fibre longitudinali appartengono ai tratti piramidali, fibrae corticopontinae, che si collegano ai nuclei propri del ponte, da dove hanno origine le fibre trasversali che vanno alla corteccia cerebellare, tractus pontocerebellaris. L'intero sistema di percorsi collega la corteccia cerebrale con la corteccia cerebellare attraverso un ponte. Quanto più sviluppata è la corteccia cerebrale, tanto più sviluppati sono il ponte e il cervelletto. Naturalmente, il ponte risulta essere più pronunciato negli esseri umani, che è una caratteristica specifica della struttura del suo cervello. Nella pars dorsalis è presente la formatio reticularis pontis, che è una continuazione della stessa parte del midollo allungato, e sopra la formazione reticolare si trova il fondo della fossa romboidale rivestita di ependima con i nuclei sottostanti dei nervi cefalici ( coppie VIII-V).
Nella pars dorsalis continuano anche i tratti conduttivi del midollo allungato, situati tra la linea mediana e il nucleo dorsale del corporis trapezoidei e parte dell'ansa mediale, lemniscus medialis; in quest'ultimo si intersecano i tratti ascendenti del midollo allungato, tractus bulbothalamicus.
Cervelletto
Cervelletto, cervelletto, è un derivato del rombencefalo, che si è sviluppato in connessione con recettori statici. Pertanto, è direttamente correlato alla coordinazione dei movimenti ed è un organo di adattamento del corpo per superare le proprietà fondamentali della massa corporea: gravità e inerzia. È anche considerato uno dei centri più alti del sistema vegetativo (simpatico) sistema nervoso(L.A. Orbeli e la sua scuola).
Lo sviluppo del cervelletto nel processo di filogenesi ha attraversato 3 fasi principali, corrispondenti ai cambiamenti nei metodi di movimento dell'animale.
Nella prima fase, quando gli animali si muovevano nell'acqua con movimenti serpentini del corpo (ciclostomi), il loro cervelletto sembrava una piccola formazione, l'auricola, che negli animali terrestri si sviluppava nel cosiddetto floccolo. Questa è la parte più antica del cervelletto: archicerebellum. Nella seconda fase, quando negli animali acquatici compaiono le pinne (pesci) e cambiano i movimenti del tronco, appare una nuova parte del cervelletto: il verme, che negli esseri umani è la parte vecchia del cervelletto, il paleocerebellum. Infine, nella terza fase, negli animali terrestri compaiono gli emisferi cerebellari, che sono la parte più nuova del cervelletto, il neocerebello, e crescono gradualmente man mano che il corpo dell'animale si allontana dal suolo e si sviluppa la locomozione con gli arti. Nell'uomo, grazie alla deambulazione eretta con l'aiuto di un paio di arti (gambe) e al miglioramento dei movimenti di presa della mano durante il travaglio, gli emisferi cerebellari raggiungono massimo sviluppo, per cui il cervelletto nell'uomo è più sviluppato che in tutti gli animali, il che costituisce una caratteristica specifica umana della sua struttura.
Il cervelletto è situato sotto i lobi occipitali degli emisferi cerebrali, dorsalmente al ponte e al midollo allungato, e si trova nella parte posteriore fossa cranica. Ha parti laterali voluminose, o emisferi, emisfero cerebellare, e quello centrale situato tra di loro parte stretta- verme, verme.
SU bordo d'attacco Il cervelletto contiene la tacca anteriore, che copre la parte adiacente del tronco encefalico. Al margine posteriore c'è una tacca posteriore più stretta che separa gli emisferi l'uno dall'altro.
La superficie del cervelletto è ricoperta da uno strato di sostanza grigia, che costituisce la corteccia cerebellare, e forma strette circonvoluzioni, folia cerebelli, separate l'una dall'altra da solchi, fessure cerebelli. Tra queste, la più profonda, fissura Horizontlis cerebelli, corre lungo il bordo posteriore del cervelletto e separa la superficie superiore degli emisferi, fdcies superiori, da quella inferiore, fdcies inferiori. Con l'aiuto di scanalature orizzontali e di altre grandi scanalature, l'intera superficie del cervelletto è divisa in una serie di lobuli, lobuli cerebelli. Tra questi è necessario evidenziare il piccolo lobo più isolato - il floccolo, che giace sulla superficie inferiore di ciascun emisfero in corrispondenza del peduncolo cerebellare medio, così come la parte del verme associata al floccolo - nodulo, nodulo. Il floccolo è collegato al nodulo mediante una sottile striscia - il peduncolo del floccolo, pedunculus flocculi, che medialmente passa in una sottile lamina semilunare - il velo midollare inferiore, velum medullare inferius.
Struttura interna del cervelletto. Nello spessore del cervelletto sono presenti nuclei accoppiati di materia grigia, situati in ciascuna metà del cervelletto, tra la sua sostanza bianca (Fig. 281). Ai lati della linea mediana, nell'area in cui la tenda, fastigio, sporge nel cervelletto, si trova il nucleo più mediale: il nucleo della tenda, nucleo fasiigii. Lateralmente ad esso si trovano piccole isole del nucleo sferico, nucleo globoso, e ancora più laterali - nucleo a forma di sughero, nucleo emboliforme. Infine, al centro dell'emisfero c'è un nucleo dentato, il nucleo denttus, che sembra una piastra tortuosa grigia, simile al nucleo dell'olivo, e i nuclei dentati dell'ilo aperti medialmente. La somiglianza tra il nucleo dentato del cervelletto e il nucleo olivare, anch'esso frastagliato, non è casuale, poiché entrambi i nuclei sono collegati da vie, fibrae olivocerebellares, e ciascuna circonvoluzione di un nucleo è simile alla circonvoluzione dell'altro. Pertanto, entrambi i nuclei partecipano insieme all'implementazione della funzione di equilibrio (vedi Fig. 280, 281).
I nuclei cerebellari nominati hanno età filogenetiche diverse: nucleo fastigii si riferisce alla parte più antica del cervelletto - flocculus (archicerebellum), associato all'apparato vestibolare, nuclei emboliformis et globosus - alla parte vecchia (paleocerebellum), sorta in connessione con i movimenti del corpo e nucleo dentato - al più giovane (neocerebello), che si è sviluppato in connessione con il movimento con l'aiuto degli arti. Pertanto, quando ciascuna di queste parti viene colpita, vengono violati aspetti diversi funzione motoria, corrispondente diverse fasi filogenesi, vale a dire: quando il sistema flocculonodulare e il suo nucleo tenda sono danneggiati, l'equilibrio del corpo è disturbato. Quando il verme e i suoi corrispondenti nuclei a forma di sughero e sferici vengono danneggiati, il funzionamento dei muscoli del collo e del busto viene interrotto. Quando sono colpiti gli emisferi e il nucleo dentato, lavorano i muscoli degli arti.
La sostanza bianca del cervelletto in una sezione ha l'aspetto di piccole foglie di una pianta, corrispondenti a ciascun giro, ricoperte alla periferia dalla corteccia della sostanza grigia. Di conseguenza, l'immagine complessiva della materia bianca e grigia su una sezione del cervelletto ricorda un albero, arbor viiae cerebelli (albero della vita; dal nome aspetto, poiché il danno al cervelletto non lo è minaccia immediata vita). La sostanza bianca del cervelletto è composta da vari tipi fibre nervose. Alcuni collegano le circonvoluzioni e i lobuli, altri vanno dalla corteccia ai nuclei interni del cervelletto e, infine, altri collegano il cervelletto con le parti vicine del cervello. Queste ultime fibre sono incluse tre paia di peduncoli cerebellari:
1. Parte inferiore delle gambe, peduncoli cerebellari inferiori(al midollo allungato).
Includono il trdctus spinocerebelldris posteriore (Flechsig) che va al cervelletto, le fibrae arcudtae externae - dai nuclei delle corde posteriori del midollo allungato e le fibrae olivocerebellar - dall'olivo. Tutte queste fibre terminano nella corteccia del verme e negli emisferi. Inoltre, ci sono fibre provenienti dai nuclei del nervo vestibolare, che terminano nel nucleo fastigii. Grazie a tutte queste fibre, il cervelletto riceve impulsi da apparato vestibolare e propriocettivo, per cui diventa il nucleo della sensibilità propriocettiva, apportando correzioni automatiche all'attività motoria di altre parti del cervello. Come parte della parte inferiore delle gambe, ci sono anche percorsi discendenti nella direzione opposta, vale a dire: dal nucleo fastigii al nucleo vestibolare laterale e da esso alle corna anteriori del midollo spinale, tractus vestibulospinalis. Attraverso questo percorso, il cervelletto influenza il midollo spinale.
2. Zampe centrali, peduncoli cerebellari medii(al ponte).
Contengono fibre nervose dai nuclei pontini alla corteccia cerebellare. Le vie di accesso alla corteccia cerebellare, trdctus pontocerebellars, che originano dai nuclei pontini, si trovano in continuazione delle vie cortico-pontine, fibrae corticopontinae, terminando nei nuclei pontini dopo la decussazione. Queste vie collegano la corteccia cerebrale con la corteccia cerebellare, il che spiega il fatto che quanto più sviluppata è la corteccia cerebrale, tanto più sviluppati sono il ponte e gli emisferi cerebellari, come si osserva negli esseri umani.
3. Parte superiore delle gambe, peduncoli cerebellari superiori(al quadrigemino). Sono costituiti da fibre nervose che corrono in entrambe le direzioni: 1) al cervelletto - trdctus spinocerebelldris anterior (Gowers), e 2) dal nucleo dentato del cervelletto al tetto del quadrigeminale - trdctus cerebellotegmentdlis, che, dopo la decussazione, termina in nel nucleo rosso e nel talamo ottico. Attraverso i primi percorsi, il cervelletto riceve impulsi dal midollo spinale e, attraverso il secondo, invia impulsi al sistema extrapiramidale, attraverso il quale esso stesso influenza il midollo spinale.
Istmo
Istmo, istmo rombencefalo, rappresenta il passaggio dal rombencefalo al mesencefalo. È costituito da: 1) i peduncoli cerebellari superiori, peduncoli cerebellari superiori; 2) il velo midollare superiore teso tra loro e il cervelletto, velo midollare superius, che è attaccato al solco mediano tra i tubercoli quadrigeminali; 3) triangolo ad anello, trigono lemnisci, causato dal decorso delle fibre uditive del lemnisco laterale, lemnisco lateralis. Questo triangolo grigio è delimitato anteriormente dal manico inferiore del quadrigemino, posteriormente dal peduncolo cerebellare superiore e lateralmente dal peduncolo cerebrale. Quest'ultimo è separato dall'istmo e dal mesencefalo da un solco chiaramente definito, sulcus lateralis mesencephali. L'estremità superiore del quarto ventricolo sporge nell'istmo e passa nell'acquedotto nel mesencefalo.
IV ventricolo
IV ventricolo, ventricuius quartus, è un residuo della cavità del rombencefalo ed è quindi una cavità comune a tutte le parti del rombencefalo che compongono il rombencefalo (midollo allungato, cervelletto, ponte e istmo). Il IV ventricolo ricorda una tenda, in cui si distinguono il fondo e il tetto (vedi Fig. 279; Fig. 282).
Metter il fondo a, o la base, del ventricolo ha la forma di un rombo, come se fosse premuto sulla superficie posteriore del midollo allungato e del ponte. Per questo lo chiamano fossa romboidale, fossa romboidea. Il canale centrale del midollo spinale si apre nell'angolo postero-inferiore della fossa romboidale, e nell'angolo antero-superiore il quarto ventricolo comunica con l'acquedotto. Gli angoli laterali terminano ciecamente sotto forma di due tasche, recessus laterdles ventriculi qudrti, curvandosi ventralmente attorno ai peduncoli cerebellari inferiori.
Tetto del IV ventricolo, tegmen ventriculi quarti, ha forma di tenda, fastigium (BNA), ed è composto da due vele cerebrali: quella superiore, velum medullare superius, tesa tra i peduncoli cerebellari superiori, e quella inferiore, velum medullare inferius, formazione paritaria adiacente al i peduncoli del cervelletto. La parte del tetto compresa tra le vele è formata dalla sostanza del cervelletto. Il velo midollare inferiore è completato da uno strato della coroide, tela chorioidea ventriculi qudrti, ricoperto dall'interno da uno strato di epitelio, lamina chorioidea epithelialis, che rappresenta un rudimento parete di fondo midollo posteriore (ad esso è associato il plesso - pl. corioideo. ventr. IV).
Il corpo corioidea inizialmente racchiude completamente la cavità del ventricolo, ma poi, durante lo sviluppo, compaiono in esso tre aperture: una nella regione dell'angolo inferiore della fossa romboidale, apertura medina ventriculi qudrti (forame Magendi) (la più grande), e due nella regione dei recessi laterali del ventricolo, aperturae laterdles ventriculi qudrti (forami Luschka). Attraverso queste aperture, il quarto ventricolo comunica con lo spazio subaracnoideo del cervello, grazie al quale liquido cerebrospinale proviene dai ventricoli cerebrali negli spazi intertecali. In caso di restringimento o fusione di questi fori a causa dell'infiammazione delle meningi (meningite), il liquido cerebrospinale che si accumula nei ventricoli cerebrali non trova sbocco nello spazio subaracnoideo e si verifica l'idropisia cerebrale.
Fossa del diamante, fossa romboidea(Fig. 283), ha quattro lati a forma di diamante: due superiori e due inferiori. I lati superiori del rombo sono delimitati dai due peduncoli cerebellari superiori, mentre i lati inferiori dai due peduncoli inferiori. Lungo il rombo, lungo la linea mediana, da angolo superiore a quello inferiore si estende un solco mediano, sulcus medidnus, che divide la fossa romboidale nelle metà destra e sinistra. Ai lati del solco si trovano una coppia di rilievi, eminenza mediale causato dall’accumulo di materia grigia.
Inferiormente l'eminentia medialis si restringe progressivamente, trasformandosi in un triangolo sul quale si proietta il nucleo del nervo ipoglosso. trigono nervi ipoglossi. Lateralmente alla parte inferiore di questo triangolo si trova un triangolo più piccolo, riconoscibile dal suo colore grigio, trigono n. vagi, che contiene il nucleo autonomo del nervo vago, nucleo dorsdlis nervi vagi. In alto, l'eminentia medialis ha un'elevazione: il tubercolo facciale, collicuius facialis, causato dal passaggio della radice facciale e dalla proiezione del nucleo del nervo abducente.
Nell'area degli angoli laterali, il campo vestibolare, area vestibularis, si trova su entrambi i lati. Qui sono posti i nuclei dell'VIII coppia. Alcune delle fibre che emergono da esse attraversano la fossa romboidale dagli angoli laterali al solco mediano sotto forma di strisce orizzontali, striae medulldres ventriculi qudrti. Queste strisce dividono la fossa romboidale in metà superiore e inferiore e corrispondono al confine tra il midollo allungato e il ponte.
Topografia della sostanza grigia della fossa romboidale. La materia grigia del midollo spinale passa direttamente nella materia grigia del tronco cerebrale e in parte si diffonde lungo la fossa romboidale e le pareti dell'acquedotto (vedi mesencefalo), e in parte è divisa in nuclei separati dei nervi cerebrali o nuclei di fasci di percorsi.
Per comprendere la posizione di questi nuclei, è necessario tenere conto, come accennato in precedenza, che il tubo midollare chiuso del midollo spinale, quando passa nel midollo allungato, si apre sul lato posteriore e si apre in una fossa romboidale. Di conseguenza, i corni posteriori della sostanza grigia del midollo spinale sembravano divergere lateralmente. I nuclei somatico-sensibili localizzati nei corni posteriori erano localizzati lateralmente nella fossa romboidale, mentre i nuclei somatico-motori corrispondenti ai corni anteriori rimanevano localizzati medialmente. Per quanto riguarda i nuclei vegetativi situati nelle corna laterali del midollo spinale, quindi, a seconda della posizione delle corna laterali tra la parte posteriore e quella anteriore, questi nuclei, quando il tubo cerebrale si espandeva, risultavano trovarsi nella fossa romboidale tra il nuclei somatico-sensibili e somatico-motori. Di conseguenza, nella parete della fossa romboidale, a differenza del midollo spinale, i nuclei della sostanza grigia non si trovano nella direzione anteroposteriore, ma si trovano in file - medialmente e lateralmente.
Quindi, ad esempio, i nuclei somatico-motori delle coppie XII e VI si trovano nella fila mediale, i nuclei vegetativi delle coppie X, IX, VII sono nella fila centrale, e i nuclei somatico-sensibili della coppia VIII sono laterale.
Proiezione dei nuclei dei nervi cerebrali sulla fossa romboidale:
XII coppia - nervo ipoglosso, n. ipoglosso, ha un unico nucleo motore situato nella parte più bassa della fossa romboidale, nella profondità del trigono n. ipoglossi.
XI coppia - nervo accessorio, n. accessorio, ha due nuclei (entrambi motori): uno è situato nel midollo spinale e si chiama nucleo spinalis n. accessorii, l'altro è una continuazione caudale dei nuclei delle coppie di nervi X e IX ed è chiamato nucleo ambiguo. Si trova nel midollo allungato dorsolaterale al nucleo olivare.
Coppia X - nervo vago, N. vago, ha tre nuclei:
1. Il nucleo sensibile, nucleo trdctus solitdrii, è situato accanto al nucleo del nervo ipoglosso, in profondità nel trigono n. Vaghi.
2. Nucleo vegetativo, nucleo dorsale i. vagi, si trova nella stessa zona.
3. Il nucleo motore, nucleo ambiguo (doppio), è in comune con il nucleo della coppia IX, situato nella formatio reticularis, più in profondità del nucleo dorsale.
IX coppia - nervo glossofaringeo, n. glossofaringeo, contiene anche tre nuclei:
1. Il nucleo sensibile, nucleo trdctus solitdrii, si trova lateralmente al nucleo del nervo ipoglosso.
2. Nucleo vegetativo (secretorio), nucleo salivatorio inferiore, nucleo salivare inferiore; le sue cellule sono sparse nella formatio reticularis del midollo allungato tra n. ambiguo e nocciolo d'oliva.
3. Nucleo motore, comune con il n. vago e n. accessorio, nucleo ambiguo.
VIII coppia - staticamente- nervo uditivo, N. vestibolococleare, ha nuclei multipli che sporgono sugli angoli laterali della fossa romboidale, nella zona vestibolare. I nuclei sono divisi in due gruppi, corrispondenti alle due parti del nervo; una parte del nervo, pars cochlearis - il nervo cocleare, o nervo uditivo stesso, ha due nuclei: quello dorsale, nucleo cocleare dorsdlis, e quello ventrale, nucleo cocleare ventrdlis, situato lateralmente e anteriormente al precedente. Un'altra parte del nervo, pars vestibularis - il nervo del vestibolo, o nervo statico, ha quattro nuclei (nuclei vestibulares):
1. Mediale - principale (nucleo Schwalbe).
2. Laterale: nucleo di Deiters.
3. Superiore: nucleo di Bekhterev.
4. In basso.
La presenza di quattro nuclei nell'uomo riflette fasi iniziali filogenesi, quando i pesci avevano diversi apparati statici percettivi separati.
VII coppia - nervo facciale, n. facciale, ha un nucleo motore situato nella formatio reticularis partis dorsalis del ponte. Le fibre nervose che si estendono da esso nel loro percorso attraverso lo spessore del ponte formano un anello che sporge sulla fossa romboidale sotto forma di collicolo facciale.
Nervo intermedio, n. intermedio, strettamente connesso nel suo decorso al nervo facciale, presenta due nuclei:
1. Autonomo (secretorio), nucleo salivatorius superiore (nucleo salivare superiore), incorporato nella formatio reticularis del ponte, dorsale al nucleo nervo facciale.
2. Sensibile, nucleo trdctus solitdrii.
VI coppia - nervo abducente, n. abdiicens, ha un nucleo motore situato nell'ansa del nervo facciale, quindi il collicolo facciale sulla superficie della fossa romboidale corrisponde a questo nucleo
Coppia V - nervo trigemino, n. trigemino, ha quattro core:
1. Sensibile, nucleo sensorius principalis n. trigemini, è proiettato nella parte dorsolaterale della parte superiore del ponte.
2. Nucleo del tratto spinale, nucleo del tratto spinale n. trigemini, è la continuazione del precedente per tutta la lunghezza del midollo allungato regione cervicale midollo spinale, dove entra in contatto con la sostanza gelatinosa delle corna posteriori.
3. Nucleo motore, nucleo motorius n. trigemini (masticatori), situati medialmente a quello sensibile.
4. Nucleo del tratto mesencefalico, nucleo del tratto mesencefalico n. trigemini, si trova lateralmente all'acquedotto. Rappresenta il nucleo della sensibilità propriocettiva per i muscoli masticatori e per i muscoli bulbo oculare.
È possibile che questo nucleo rifletta lo sviluppo autonomo del primo ramo nervo trigemino(n. ophthalmicus), chiamato n. negli animali. ophthalmicus profundus e correlato all'organo della vista, il che spiega la posizione del nucleo nel mesencefalo.
Mesencefalo
Mesencefalo, mesencefalo, si sviluppa nel processo di filogenesi sotto l'influenza predominante recettore visivo, quindi, le sue formazioni più importanti sono legate all'innervazione dell'occhio. Qui si formarono i centri dell'udito che, insieme ai centri della vista, in seguito crebbero sotto forma di quattro tubercoli: il quadrigemino. Con l'avvento dell'udito e analizzatori visivi nella corteccia proencefalo gli stessi centri uditivi e visivi del mesencefalo caddero in una posizione subordinata e divennero intermedi, sottocorticali. Con lo sviluppo del cervello anteriore nei mammiferi superiori e nell'uomo, i percorsi iniziarono a passare attraverso il mesencefalo, collegando la corteccia del midollo terminale con il midollo spinale (peduncoli cerebrali) (vedi Fig. 277, 278).
Di conseguenza, il mesencefalo umano contiene: 1) centri visivi sottocorticali e nuclei nervosi che innervano i muscoli oculari; 2) sottocorticale centri uditivi; 3) tutte le vie ascendenti e discendenti che collegano la corteccia cerebrale con il midollo spinale e passano attraverso il mesencefalo; 4) fasci di sostanza bianca che collegano il mesencefalo con altre parti del sistema nervoso centrale. Di conseguenza, il mesencefalo, che è il più piccolo e il più semplice organizzato dal dipartimento Il cervello è composto da due parti principali: il tetto, dove si trovano i centri sottocorticali dell'udito e della vista, e i peduncoli del cervello, dove passano principalmente le vie conduttive.
1. Parte dorsale, tetto del mesencefalo, O placca quadrigeminale, tectum mesencephali s. lamina quadrigemina(BNA) (vedi Fig. 278).
È nascosto sotto l'estremità posteriore del corpo calloso ed è diviso da due solchi incrociati - longitudinale e trasversale - in quattro tubercoli bianchi, disposti a coppie.
I due tubercoli superiori, colliculi superiori, sono centri di visione sottocorticali; entrambi quelli inferiori, colliculi inferiores, sono centri dell'udito sottocorticali. In un solco piatto tra i tubercoli superiori si trova ghiandola pineale. Ciascun tubercolo passa nel cosiddetto ansa del tubercolo, brachium colliculi, diretto lateralmente, anteriormente e verso l'alto, verso diencefalo. L'ansa del tubercolo superiore, brdchium collculi superioris, va sotto il cuscino, pulvinar, del tubercolo ottico al corpo genicolato laterale, corpus geniculatum laterale. Il manico del tubercolo inferiore, brdchium collculi inferioris, che passa lungo il bordo superiore del trigonum lemnisci fino al solco laterale mesencefali, scompare sotto il corpo genicolato mediale, corpus geniculatum mediale. I corpi genicolati menzionati appartengono già al diencefalo.
2. Parte ventrale, peduncoli del cervello, peduncoli cerebri, contiene tutti i percorsi per il prosencefalo.
I peduncoli cerebrali sembrano due spesse corde bianche semicilindriche che divergono ad angolo dal bordo del ponte e si tuffano nello spessore degli emisferi cerebrali.
3. La cavità del mesencefalo, che è un residuo della cavità primaria della vescica del mesencefalo, ha la forma canale stretto e viene chiamato acqua corrente,acquedotto cerebrale(Silvii). È uno stretto canale rivestito ependimale, lungo 1,5-2,0 cm, che collega il ventricolo IV con il III. Dorsalmente l'acquedotto è limitato dal tetto del mesencefalo e ventralmente dal tegmento dei peduncoli cerebrali.
Struttura interna del mesencefalo. In una sezione trasversale del mesencefalo si distinguono tre parti principali del tronco cerebrale: 1) la piastra del tetto, lamina tetto, formato dal quadrigemino; 2) pneumatico, tegmento, che rappresentano sezione superiore peduncoli cerebrali; 3) la sezione ventrale dei peduncoli cerebrali o il peduncolo cerebrale stesso, crus cerebrale. Secondo lo sviluppo del mesencefalo sotto l'influenza del recettore visivo (E.K. Sepp), contiene vari nuclei legati all'innervazione dell'occhio.
Nei vertebrati inferiori, il collicolo superiore funge da terminazione principale del nervo ottico ed è il centro visivo. Nei mammiferi e nell'uomo, con il trasferimento dei centri visivi al prosencefalo, la restante connessione del nervo ottico con il collicolo superiore è importante solo per i riflessi. Le fibre dell'ansa uditiva (lemniscus lateralis) terminano nel nucleo del collicolo inferiore, così come nel corpo genicolato mediale. Il tetto del quadrigemino ha un collegamento bidirezionale midollo spinale- trdctus spinotecdlis e trdctus tectobulbdris et tectospindlis. Questi ultimi, dopo la decussazione nel tegmento (decussazione dorsale a fontana di Meynert), si dirigono verso i nuclei muscolari del midollo allungato e del midollo spinale. Questa è la cosiddetta via del riflesso visivo-sonoro, di cui abbiamo parlato nella descrizione del midollo spinale. Pertanto, la regione quadrigeminale può essere considerata un centro riflesso per vari tipi di movimenti che si verificano principalmente sotto l'influenza di stimoli visivi e uditivi.
L'acquedotto silviano è circondato da un acquedotto centrale materia grigia, che nella sua funzione è correlato a sistema autonomo. In esso, sotto la parete ventrale dell'acquedotto, nel tegmento del peduncolo cerebrale, si trovano i nuclei di due nervi motori del cervello: N. oculomotorio(III paio) a livello del collicolo superiore e N. trocleare(IV paio) a livello del collicolo inferiore. Nucleo nervo oculomotoreè costituito da diverse sezioni, corrispondenti all'innervazione di diversi muscoli del bulbo oculare. Medialmente e posteriormente ad esso si trova un altro piccolo nucleo vegetativo, anch'esso pari, nucleo accessorio oppure il nucleo Yakubovich (Yakubovich lo descrisse nel 1857, prima di Westphal ed Edinger, da cui venne erroneamente chiamato) e il nucleo mediano spaiato. Il nucleo Yakubovich e il nucleo mediano spaiato innervano i muscoli lisci dell'occhio, m. ciliare e m. pupille dello sfintere. Questa parte del nervo oculomotore appartiene sistema parasimpatico. Sopra (orale) il nucleo del nervo oculomotore nel tegmento del peduncolo cerebrale si trova il nucleo del fascicolo longitudinale mediale (nucleo di Darkshevich).
Lateralmente all'acquedotto si trova il nucleo del tratto mesencefalico del nervo trigemino, nucleo del tratto mesencefalico n. trigemini.
I peduncoli cerebrali sono divisi, come già accennato, nella parte ventrale, o il vero e proprio peduncolo del cervello, crus cerebri, E pneumatico, tegmento. Il confine tra loro è la sostanza nera, substantia nigra, che deve il suo colore al pigmento nero contenuto nelle sue cellule nervose costituenti: la melanina (Fig. 284).
Sostantia nigra si estende lungo tutta la lunghezza del peduncolo cerebrale dal ponte al diencefalo; per la sua funzione appartiene al sistema extrapiramidale.
Situato ventralmente alla substantia nigra, il crus cerebri contiene fibre nervose longitudinali che scendono dalla corteccia cerebrale a tutte le parti sottostanti del sistema nervoso centrale (tratto corticopontino, corticonuclearis, corticospinale, ecc.). Il tegmento, situato dorsalmente alla substantia nigra, contiene principalmente varie fibre ascendenti, oltre a nuclei di sostanza grigia. Tra questi, il più significativo è nucleo rubino.
Questa formazione allungata a forma di salsiccia si estende nel tegmento del peduncolo cerebrale dalla regione subtubercolare del diencefalo al collicolo inferiore, dove inizia un importante tratto discendente, tractus rubrospinalis, che collega il nucleo rosso con le corna anteriori del midollo spinale. Questo fascio, dopo aver lasciato il nucleo rosso, si interseca con un fascio simile del lato opposto nella parte ventrale della sutura mediana - la decussazione ventrale del tegmento (Trota). Il nucleo ruber è un centro di coordinamento molto importante del sistema extrapiramidale, collegato con le altre sue parti. Le fibre gli passano dal cervelletto come parte dei peduncoli superiori di quest'ultimo dopo la loro decussazione sotto il quadrigemino, ventralmente dall'acquedotto cerebrale, così come dal pallido - questo è il più basso e il più antico dei nodi sottocorticali del cervello che fanno parte del sistema extrapiramidale. Grazie a queste connessioni, il cervelletto ed il sistema extrapiramidale, attraverso il nucleo rosso ed il tratto rubrospinale che da esso si estende, influenzano l'intero muscoli scheletrici nel senso di movimenti automatici inconsci. Oltre alle fibre longitudinali discendenti, il tegmento contiene fibre ascendenti, che formano una continuazione delle anse mediale (lemniscus medialis) e laterale (lemniscus lateralis) nel mesencefalo. Come parte di questi anelli, tutti ascendono al cervello percorsi sensoriali, ad eccezione di quello visivo e olfattivo.
Continua anche il tegmento del mesencefalo formazione reticolare, formatio reticularis, E fasciculus longitudindlis medialis. Quest'ultimo ha origine in vari luoghi. Una delle sue parti inizia da nuclei vestibolari, passa su entrambi i lati lungo i lati della linea mediana, direttamente sotto la materia grigia del fondo dell'acquedotto silviano e del IV ventricolo, ed è costituito da fibre ascendenti e discendenti che vanno a nuclei III, IV, VI e XI nervi cefalici. Fascicolo longitudinale medialeè un'importante via di associazione che collega vari nuclei nervosi muscoli oculari tra loro, che determina i movimenti combinati degli occhi quando deviano in una direzione o nell'altra. La sua funzione è associata anche ai movimenti degli occhi e della testa che avvengono quando viene stimolato l'apparato dell'equilibrio.
Prosencefalo
Proencefalo, prosencefalo, si sviluppa in connessione con il recettore olfattivo e inizialmente (negli animali acquatici) è un cervello puramente olfattivo, il rinencefalo. Con il passaggio degli animali dall'ambiente acquatico all'aria, aumenta il ruolo del recettore olfattivo, poiché con il suo aiuto determinano le sostanze contenute nell'aria. sostanze chimiche, segnalando all'animale prede, pericoli e altri elementi vitali eventi importanti la natura da una lunga distanza: un recettore distante.
Pertanto, e anche grazie allo sviluppo e al miglioramento di altri analizzatori, il cervello anteriore degli animali terrestri cresce notevolmente e supera altre parti del sistema nervoso centrale, trasformandosi da cervello olfattivo in un organo che controlla l'intero comportamento dell'animale. Secondo due principali forme di comportamento: 1) comportamento istintivo basato sull'esperienza della specie ( senza riflessi condizionati), e 2) comportamento individuale basato sull'esperienza dell'individuo (riflessi condizionati) - due gruppi di centri si sviluppano nel prosencefalo: 1) nodi basali o centrali degli emisferi cerebrali ("sottocorteccia"); 2) corteccia cerebrale. Questi due gruppi di centri del proencefalo ricevono tutti gli impulsi nervosi e ad essi si estendono tutte le vie sensoriali afferenti, che (con poche eccezioni) passano prima attraverso un centro comune: il talamo visivo, o camera, talamo. L'adattamento dell'organismo all'ambiente attraverso i cambiamenti del metabolismo ha portato alla comparsa nel prosencefalo di centri superiori responsabili dei processi vegetativi (regione subtalamica, ipotalamo).
Delle due parti del prosencefalo, il diencefalo, diencefalo e finale telencefalo, la corteccia e i nodi sottocorticali appartengono al telencefalo, mentre il talamo visivo e la regione subtalamica appartengono al cervello intermedio. Secondo E.K. Sepp (1959), il diencefalo si è sviluppato nel processo di filogenesi non dal prosencefalo come risultato della divisione di quest'ultimo, ma insieme al mesencefalo. Ciò spiega la connessione del recettore visivo non solo con il mesencefalo, ma anche con il cervello intermedio e il suo talamo visivo, che quindi ha ricevuto il nome corrispondente.
Diencefalo
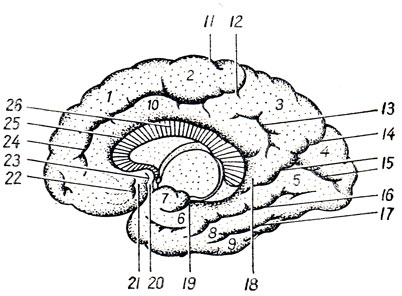
Diencefalo, si trova sotto il corpo calloso e il fornice, fuso lateralmente con gli emisferi del telencefalo. Secondo quanto detto sopra circa la funzione e lo sviluppo del prosencefalo, nel diencefalo si distinguono due parti principali: 1) dorsale (filogeneticamente più giovane) - talamencefalo- centro delle vie afferenti e 2) ventrale (filogeneticamente più antico) - ipotalamo- il centro vegetativo più alto. La cavità del diencefalo è III ventricolo.
Talamomencefalo
Talamomencefalo a sua volta, è costituito da tre parti: talamo - il palazzo, epitalamo - la regione sopratalamica e metatalamo - la regione transtalamica (Fig. 285).
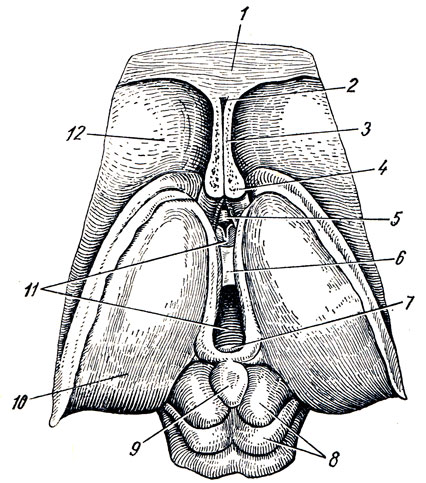
UN. Talamo, talamo, è un grosso accumulo appaiato di materia grigia nelle pareti laterali del diencefalo ai lati del terzo ventricolo, di forma ovoidale, con l'estremità anteriore appuntita a forma di tuberculum anterius, e l'estremità posteriore espansa e ispessita in la forma di un cuscino, pulvinar. La divisione nell'estremità anteriore e nel cuscino corrisponde alla divisione funzionale del talamo nei centri delle vie afferenti (estremità anteriore) e nel centro visivo (posteriore). La superficie dorsale è coperta strato sottile sostanza bianca - strato zonule. Nella sua sezione laterale è rivolto verso la cavità ventricolo laterale, separato dall'adiacente nucleo caudato da un solco di confine, sulcus termindlis, che costituisce il confine tra il telencefalo, a cui appartiene il nucleo caudato, e il diencefalo, a cui appartiene il talamo ottico. Una striscia di midollo, stria termindlis, corre lungo questo solco.
La superficie mediale del talamo visivo, ricoperta da uno strato di sostanza grigia centrale, è verticale e si affaccia sulla cavità del terzo ventricolo, formandone la parete laterale. Dall'alto è delimitato dalla superficie dorsale dalla striscia midollare bianca, stria medulldris thalami. Entrambe le superfici mediali delle tuberosità visive sono collegate tra loro da una commessura grigia - adhesio interthaldmica, che giace quasi al centro. La superficie laterale del talamo visivo confina con la borsa interna, la capsula interna. Con la sua superficie inferiore, il talamo visivo si trova sopra il peduncolo cerebrale, fondendosi con il suo opercolo (Fig. 286). Come si può vedere nelle sezioni, la massa grigia del talamo visivo è divisa da strati bianchi, lamine medullares talami, in nuclei separati, denominati a seconda della loro topografia: anteriore, centrale, mediale, laterale e alcuni ventrali.
Significato funzionale il talamo visivo è molto grande. In esso si scambiano vie afferenti: nel suo cuscino, pulvinar, parte delle fibre del tratto ottico termina (centro della visione sottocorticale), in nucleo anteriore- Fascio di Vic d'Azir, proveniente dai corpi mamillaria e che collega il tubercolo visivo con la sfera olfattiva, e, infine, tutte le altre vie sensoriali afferenti dalle parti sottostanti del sistema nervoso centrale nei suoi restanti nuclei, con terminazione del lemnisco mediale nel nucleo laterale. Pertanto il talamo è il centro sottocorticale di tutti i tipi di sensibilità. Da qui le vie sensoriali vanno in parte ai nodi sottocorticali (per cui il talamo ottico diventa il centro sensibile del sistema extrapiramidale), in parte direttamente al corteccia (tratto talamocorticale).
B. Epitalamo. Le strie midollari di entrambe le tuberosità visive sono dirette posteriormente (caudalmente) e formano un'estensione triangolare su entrambi i lati, chiamata trigonum habenulae. Da quest'ultimo deriva il cosiddetto guinzaglio, habenula, che, insieme allo stesso guinzaglio del lato opposto, si collega al corpo pineale, corpus pineale (pinus - pine) s. epifisi (vedere Fig. 285). Davanti al corpo pineale entrambi i guinzagli sono legati insieme mediante la commissura habenuldrum. Il corpo pineale stesso, somigliante a diversi pigna(da cui il nome), per struttura e funzione appartiene alle ghiandole secrezione interna. Il corpo pineale, sporgendo posteriormente nella regione del mesencefalo, si trova nel solco tra i collicoli superiori dei quadrigemini, formando, per così dire, il quinto tubercolo.
IN. Metatalamo. Dietro il talamo visivo si trovano due piccoli rilievi: i corpi genicolati, corpus geniculdtum laterale et mediato (vedi Fig. 278, 286).
Il corpo genicolato mediale, di dimensioni più piccole ma più pronunciato, giace davanti all'ansa inferiore della quadrigemina sotto il pulvinar del talamo ottico, separato da esso da un evidente solco. Le fibre dell'anello uditivo, lemnisco laterale, terminano in esso, per cui, insieme ai tubercoli inferiori del quadrigemino, costituisce il centro sottocorticale dell'udito. Il corpo genicolato laterale, più grande, a forma di tubercolo piatto, è situato sulla faccia laterale inferiore del pulvinar. In esso termina per la maggior parte la parte laterale del tratto ottico (l'altra parte del tratto termina nel pulvinar). Pertanto, insieme al pulvinar e al collicolo superiore, il corpo genicolato laterale costituisce il centro della visione sottocorticale. I nuclei di entrambi i corpi genicolati sono collegati tramite vie centrali alle estremità corticali dei corrispondenti analizzatori.
Ipotalamo
Regione subtalamica, ipotalamo, nel senso lato del termine, unisce formazioni situate ventralmente sotto il fondo del terzo ventricolo, davanti alla substantia perforata posterior, comprendente la regione subtubercolare in senso stretto, regio subthalamica. Rispettivamente sviluppo embrionale l'ipotalamo è diviso in due sezioni: anteriore - visivo, pars optica Hypothalami, sotto il nome del quale tuber cinereum è combinato con infundibolo e ipofisi, nonché chiasma ottico con tractus opticus e posteriore - olfattivo: corpi mamillaria e regio subthalamica.
UN. Tuber cinereum, tubercolo grigio, situato davanti ai corpi mamillaria, è una sporgenza cava spaiata della parete inferiore del terzo ventricolo, costituita da una sottile lamina di sostanza grigia. L'apice del tubercolo è allungato in uno stretto imbuto cavo, l'infundibolo, alla cui estremità cieca si trova un'appendice cerebrale, ipofisi(glandula pituitaria), giacente nel recesso della sella turcica (per la sua descrizione vedi la sezione “Organi interni di secrezione”). Il tuber cinereum contiene i nuclei della materia grigia, che sono i centri vegetativi più alti, che influenzano, in particolare, il metabolismo e la termoregolazione.
B. Chiasma ottico, chiasma ottico, si trova davanti al tubercolo grigio, formato da una decussazione nervi ottici, nn. ottica.
IN. Corpi mamillaria, corpi mastoidi, due piccoli rilievi bianchi di forma irregolarmente sferica, giacenti simmetricamente ai lati della linea mediana, davanti alla substantia perforata posteriore. Sotto lo strato superficiale di sostanza bianca, all'interno di ciascuno dei corpi si trovano due nuclei grigi.
Secondo la loro funzione, i corpi mamillaria appartengono alla sottocorticale centri olfattivi.
G. Regio subthalamica, regione sottocutanea(nel senso stretto del termine); Questa è una piccola area del midollo situata sotto il talamo ottico. In esso, lateralmente alla substantia nigra, si trova il diencefalo corpo ovale, corpus subthalamicum Luysi. Il corpo di Lewis è uno degli anelli del sistema extrapiramidale; gli viene anche attribuito il merito funzioni autonome.
III ventricolo
III ventricolo, ventriculus tertlus, si trova proprio lungo la linea mediana e sulla sezione frontale del cervello si presenta come una stretta fessura verticale. Le pareti laterali del terzo ventricolo sono formate dalle superfici mediali delle tuberosità visive, tra le quali si estende quasi al centro l'adesio interthalamica (vedi Fig. 285). Parete anteriore Il ventricolo è composto inferiormente da una lamina sottile, la lamina terminalis, e più in alto da colonne del fornice (columnae fornicis) attraversate da una commissura anteriore bianca, commissura cerebri anterior. Ai lati della parete anteriore del ventricolo, le colonne del fornice, insieme alle estremità anteriori delle tuberosità visive, delimitano i fori interventricolari, forami interventricularia (Monroi), collegando la cavità del terzo ventricolo con i ventricoli laterali situati negli emisferi del telencefalo. Parete superiore Il terzo ventricolo, situato sotto il fornice e il corpo calloso, è la tela chorioidea ventriculi tertii (Fig. 287), quest'ultima comprende la parete sottosviluppata della vescica cerebrale sotto forma di placca epiteliale, lamina epiteliale e fusa con essa coroide. Ai lati della linea mediana nella tela corioidea è presente un plesso villoso, il plesso chorioideus ventriculi tertii. In zona parete di fondo del ventricolo vi sono la commissura habeniilarum et commissura cerebri posterior, tra le quali la protuberanza cieca del ventricolo, recessus pinealis, sporge nel lato caudale. Ventralmente dalla commissura posteriore, l'acquedotto si apre nel terzo ventricolo con un'apertura a forma di imbuto. Parete inferiore stretta Il terzo ventricolo, delimitato dall'interno dalle pareti laterali da solchi (sulci ipotalamici), dal lato della base del cervello corrisponde alla substantia perforata posterior, corpora marnillaria, tuber cinereum con chiasma opticum. Nella zona del fondo, la cavità ventricolare forma due depressioni: il recessus infundibuli, sporgente nel tubercolo grigio e nell'imbuto, e il recessus opticus, situato anteriormente al chiasma. La superficie interna delle pareti del terzo ventricolo è ricoperta di ependima.
Le parti del cervello considerate: romboidale (ad eccezione del cervelletto), media e intermedia sono combinate sotto il nome tronco encefalico. I medici a volte si riferiscono solo al rombencefalo e al mesencefalo con il tronco encefalico (E.K. Sepp, M.B. Zucker, E.V. Schmidt, 1950). Il tronco cerebrale, una formazione filogeneticamente più antica, differisce significativamente nella struttura e nella funzione dalla parte più giovane del cervello, il telencefalo.
telencefalo
Come menzionato sopra, telencefalo, rappresentato da due emisferi, emisfero cerebrale. Ciascun emisfero comprende: un mantello, o manto, pallio, cervello olfattivo, rinencefalo e nodi della base. Il resto delle cavità originali di entrambe le vescicole del telencefalo sono i ventricoli laterali, ventriculi lateralis. Il proencefalo, da cui viene rilasciato il telencefalo, appare prima in connessione con il recettore olfattivo (cervello olfattivo), e poi diventa l'organo di controllo del comportamento dell'animale, e in esso sorgono centri di comportamento istintivo basati sulle reazioni della specie (riflessi incondizionati). esso - nodi sottocorticali e centri di comportamento individuale basati sull'esperienza individuale (riflessi condizionati) - la corteccia cerebrale. Pertanto nel telencefalo si distinguono, in ordine di sviluppo storico: gruppi di centri:
1. Il cervello olfattivo, il rinencefalo, è la parte più antica e allo stesso tempo più piccola, situata ventralmente.
2. Nodi basali, o centrali, degli emisferi, “sottocorteccia”, la parte vecchia del telencefalo, paleencefalo, nascosta nelle profondità.
3. La materia grigia della corteccia, corteccia, la parte più giovane, neencefalo, e allo stesso tempo la parte più grande, che copre il resto come se fosse un mantello, da qui il suo nome: mantello, o mantello, pallio.
Oltre alle due forme di comportamento note per gli animali, negli esseri umani si presenta una terza forma: il comportamento collettivo, basato sull'esperienza del collettivo umano, creato nel processo dell'attività lavorativa umana e della comunicazione tra le persone attraverso la parola. Questa forma di comportamento è associata allo sviluppo degli strati superficiali più giovani corteccia cerebrale, costituendo il substrato materiale del cosiddetto secondo sistema di segnale (verbale) della realtà (I. P. Pavlov).
Poiché nel processo di evoluzione, tra tutte le parti del sistema nervoso centrale, il telencefalo cresce più velocemente e più forte, diventa la parte più per la maggior parte cervello e assume la forma di due voluminosi emisferi: destro e sinistro, hemispherium dextrum et sinistrum. Nelle profondità della fessura longitudinale del cervello, entrambi gli emisferi sono collegati tra loro da una spessa piastra orizzontale - corpo calloso, corpo calloso, che consiste di fibre nervose che corrono trasversalmente da un emisfero all'altro. Nel corpo calloso vi è un'estremità anteriore piegata verso il basso, o ginocchio, genu corporis calli, una parte mediana, truncus corporis calli, e poi un'estremità posteriore, ingrossata a forma di rullo, splenium corporis calli. Tutte queste parti sono chiaramente visibili in una sezione longitudinale del cervello tra i due emisferi (vedi Fig. 282). Il ginocchio del corpo calloso, curvandosi verso il basso, si acuisce e forma un becco, rostrum corporis callosi, che passa in una lamina sottile, lamina rostralis, che a sua volta continua nella lamina terminalis.
Sotto il corpo calloso si trova il cosiddetto volta, fornice(vedi Fig. 282; Fig. 288), raffiguranti due cordoni bianchi arcuati, che nella loro parte mediana, corpo fornico, sono collegati tra loro e divergono davanti e dietro, formando colonne ad arco davanti, Сolumnae fornicis, dietro ci sono le gambe dell'arco, crura fornicis. Crura fornicis, dirigendosi all'indietro, discende nei corni inferiori dei ventricoli laterali e lì passa nella fimbria ippocampi. Tra la crura fornicis, sotto lo splenium corporis callosi, si estendono fasci trasversali di fibre nervose, formando la commissura fornicis. Le estremità anteriori del fornice, columnae fornicis, continuano fino alla base del cervello, dove terminano nei corpi mamillares, passando attraverso la materia grigia dell'ipotalamo. Le Columnae fornicis sono limitate dai fori interventricolari che si trovano dietro di esse, collegando il terzo ventricolo con i ventricoli laterali. Davanti alle colonne dell'arco c'è una commissura anteriore, commissura anteriore, che sembra una barra trasversale bianca costituita da fibre nervose. Una sottile placca verticale è tesa tra la parte anteriore dell'arco e il genu corporis callosi tessuto cerebrale - partizione trasparente, septum pellucidum, nel cui spessore è presente una piccola cavità a fessura, cavum septi pellucidi (vedi Fig. 282, 293).
Riso. 288. Fornice, ippocampo e commissura anteriore. 1 - corpo fornico; 2 - cm fornicis; 3 - fimbria ippocampi; 4, 5 - colonna fornicis; 6 - corpo mamillare; 7 - fasciculus thalamomamillaris (Vicq - d "Azyr); 8, 9 - comissura anteriore; 10 - splenium corporis callosi; 11 - trigonum collaterale; 12 - calcar avis; 13 - polus occipitalis; 14 - polus temporalis; 15 - gyrus temporalis medius ; 16 - ippocampo
Per comodità di studio, iniziamo la descrizione delle parti in ordine inverso. sviluppo storico, cioè da un mantello che copre le restanti parti.
Pallio
In ciascun emisfero si possono distinguere tre superfici: superolaterale, mediale e inferiore, e tre bordi: superiore, inferiore e mediale, tre estremità, o poli: polo anteriore, polus frontalis, posteriore, polus occipitalis, e poi polus temporalis, corrispondenti a la sporgenza della superficie inferiore e separata su di essa da una fossa, fossa lateralis cerebri (Sylvii).
Superficie dell'emisfero (mantello), è formato da uno strato uniforme di sostanza grigia, spesso 1,3-4,5 mm, contenente cellule nervose. Questo strato, chiamato anche corteccia cerebrale, corteccia cerebrale, sembra piegato, per cui la superficie del mantello ha a massimo grado disegno complesso, costituito da solchi alternati in direzioni diverse e creste tra di loro, chiamate convoluzioni*, giri. La dimensione e la forma delle scanalature sono soggette a significative fluttuazioni individuali, a seguito delle quali non solo il cervello persone diverse, ma anche gli emisferi dello stesso individuo non sono del tutto simili nella disposizione dei solchi (vedi classificazione dei solchi).
* (L'area totale della corteccia umana adulta è di circa 220.000 mm 2 (Geneberg), di cui 2/3 si trovano in profondità tra i giri e solo 1/3 si trovano in superficie.)
Scanalature profonde e permanenti vengono utilizzate per dividere ciascun emisfero in grandi aree chiamate lobi. lobi; questi ultimi, a loro volta, sono suddivisi in lobuli e convoluzioni. Ci sono cinque lobi in ciascun emisfero: frontale ( lobo frontale), parietale ( lobo parietale), temporale ( lobo temporale), occipitale ( lobo occipitale) e un lobulo nascosto sul fondo della fessura laterale (Silviana), il cosiddetto isolotto ( insula).
Superficie superolaterale l'emisfero è delimitato in lobi da tre solchi (vedi Fig. 274; Fig. 289): le estremità laterale, centrale e superiore del solco parieto-occipitale, che, trovandosi sul lato mediale dell'emisfero, forma una tacca sul suo bordo superiore. Solco laterale, solco cerebrale laterale(Sylvii), inizia sulla superficie basale dell'emisfero dalla fossa silviana e poi passa alla superficie superolaterale, andando all'indietro e leggermente verso l'alto. Termina approssimativamente al confine del terzo medio e posteriore della superficie superolaterale dell'emisfero. Nella parte anteriore del solco laterale si dipartono due piccoli rami: uno, ramo ascendente, e l'altro, ramo anteriore, diretto al lobo frontale.
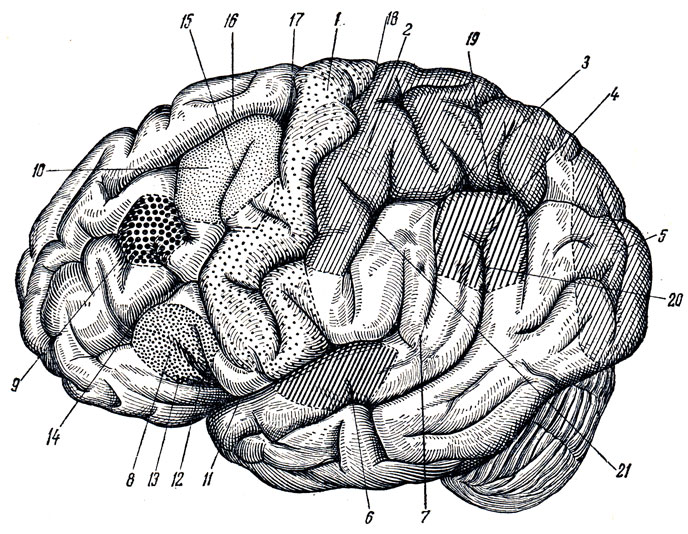
Centrale, O Rolandova, solco, solco centrale(Rolandi), inizia sul bordo superiore dell'emisfero, un po' posteriormente al suo centro, e va in avanti e in basso. L'estremità inferiore del solco centrale non raggiunge il solco laterale. La parte dell'emisfero situata davanti al solco centrale appartiene al lobo frontale; la parte della superficie cerebrale situata posteriormente al solco centrale costituisce il lobo parietale, che è delimitato dalla parte posteriore del solco laterale dal lobo temporale sottostante. Il bordo posteriore del lobo parietale è l'estremità del suddetto solco parieto-occipitale, solco parietooccipitale, situato sulla superficie mediale dell'emisfero, ma questo confine è incompleto, perché il solco nominato non si estende molto fino alla superficie superolaterale, per cui il lobo parietale passa direttamente nel lobo occipitale. Anche quest'ultimo non ha un confine netto che lo separi dal lobo temporale anteriore. Di conseguenza, il confine tra i lobi appena menzionati viene tracciato artificialmente per mezzo di una linea che va dal solco parieto-occipitale al bordo inferiore dell'emisfero.
Ciascun lobo è costituito da una serie di circonvoluzioni, talvolta chiamate lobuli, delimitate da solchi sulla superficie del cervello.
Lobo frontale. Nella regione posteriore superficie esterna questa quota passa solco precentrale quasi in direzione parallela al solco centrale. Da esso si estendono due scanalature in direzione longitudinale: solco frontale superiore et solco frontale inferiore. In tal modo Lobo frontaleè diviso in quattro circonvoluzioni: una verticale e tre orizzontali. giro verticale, giro precentrale, si trova tra il sulcus centralis e il sulcus precentralis.
Orizzontale circonvoluzioni del lobo frontale i seguenti: 1) frontale superiore, giro frontale superiore, che supera il solco frontalis superiore, parallelo al bordo superiore dell'emisfero, raggiungendone la superficie mediale; 2) giro frontale medio, giro frontale medio, si estende tra i solchi frontali superiore e inferiore e 3) giro frontale inferiore, giro frontale inferiore, posto tra il solco frontale inferiore e il solco laterale. I rami del solco laterale, estendendosi nel giro frontale inferiore, dividono quest'ultimo in tre parti: pars opercularis, situato tra l'estremità inferiore del solco precentralis e ramo ascendens sulci lateralis, pars triangularis, situato tra entrambi i rami del solco laterale , e, infine, pars orbitalis, situato di fronte al ramo anteriore del solco laterale.
Lobo parietale. Su di esso si trova approssimativamente parallelo alla scanalatura centrale solco postcentrale, di solito fondendosi con solco intraparietale, andando in direzione orizzontale. A seconda della posizione di questi solchi, il lobo parietale è diviso in tre giri, di cui uno verticale e gli altri due orizzontali. giro verticale, giro postcentrale, va dietro il solco centrale nella stessa direzione del giro precentrale, separato da esso solco centrale. Sopra il solco intraparietalis si trova il giro parietale superiore, o lobulo, lobulo parietale superiore, che si estende alla superficie mediale dell'emisfero. Al di sotto si trova il solco intraparietalis lobulo parietale inferiore, che, tornando indietro, aggira le estremità del solco laterale e del solco temporale superiore e si perde nella zona Lobo occipitale. La parte del lobulo parietale inferiore che circonda il solco laterale è chiamata giro supramargindlis; l'altra parte, che circonda il solco temporale superiore, è chiamata gyrus anguldris.
Lobo temporale. La superficie laterale di questo lobo presenta tre giri longitudinali, delimitati l'uno dall'altro solco temporale superiore E solco temporale inferiore. La parte superiore delle convoluzioni, giro temporale superiore, situato tra il solco laterale e il solco temporale superiore. La sua superficie superiore, nascosta nella profondità del solco laterale, reca 2-3 brevi circonvoluzioni chiamate gyri tempordles transversi (convoluzioni di Heschl). si estende tra i solchi temporali superiore e inferiore giro temporale medio. Al di sotto di quest'ultimo, separando da esso, passa il solco temporale inferiore giro temporale inferiore, che attraverso bordo inferiore separato dal giro occipitotemporalis lateralis che giace sulla superficie inferiore.
Lobo occipitale. I solchi sulla superficie laterale di questo lobo sono variabili e inconsistenti. Di questi si distingue quello trasversale solco occipitale trasverso, solitamente collegato all'estremità del solco intraparietalis.
Isolotto, insula. Per vedere questo lobulo è necessario allontanare o rimuovere i bordi della scanalatura laterale che lo sovrasta. Questi bordi, appartenenti ai lobi frontale, parietale e temporale, sono chiamati tegmento, opercolo. L'isola ha la forma di un triangolo, la cui parte superiore è rivolta in avanti e verso il basso. Davanti, sopra e dietro l'isola è delimitata dalle parti vicine da un profondo solco, solco circolare. La superficie dell'insula è ricoperta da brevi convoluzioni.
Superficie inferiore dell'emisfero(vedi Fig. 276) in quella parte di essa che si trova anteriormente alla fossa silviana, appartiene al lobo frontale. Qui corre parallelo al bordo mediale dell'emisfero solco olfattivo, in cui giacciono bulbus et tractus olfactorius. Un giro rettilineo si estende tra questo solco e il bordo mediale dell'emisfero, giro retto, che è una continuazione del giro frontale superiore. Lateralmente al solco olfattivo sulla superficie inferiore sono presenti numerosi solchi irregolari, solchi orbitali, limitante giri orbitali, che può essere considerato come una continuazione del giro frontale medio e inferiore. Si forma la porzione posteriore della superficie basale dell'emisfero superficie inferiore lobi temporali e occipitali, che qui non hanno confini definiti. In quest'area sono visibili due solchi: solco occipitotemporalis, passando nella direzione dal polo occipitale a quello temporale e limitante giro occipitotemporale laterale e correndo parallelo ad esso collaterali del solco(la sua continuazione anteriore è il solco rhindlis). Tra di loro si trova giro occipitotemporale mediale. Medialmente dal solco collateralis ci sono due giri: tra sezione posteriore si trova questo solco e il solco calcarinus giro linguale; tra la sezione anteriore di questo solco e il solco rinale da un lato e il profondo solco ippocampale, che circonda il tronco encefalico, dall'altro si trova giro paraippocampale. Questo giro, adiacente al tronco encefalico, si trova già sulla superficie mediale dell'emisfero.
Superficie mediale dell'emisfero. Su questa superficie (vedi Fig. 282) è presente un solco del corpo calloso, sulcus corporis callosi, che corre direttamente sopra il corpo calloso e prosegue con la sua estremità posteriore in profondità solco dell'ippocampo, che è diretto in avanti e verso il basso. Parallelamente e sopra questo solco corre lungo la superficie mediale dell'emisfero solco cingolato, che inizia davanti sotto il becco del corpo calloso, poi risale e termina con la sua estremità posteriore sul bordo superiore dell'emisfero. Lo spazio situato tra questo bordo dell'emisfero e il solco cinguli appartiene al lobo frontale, alla circonvoluzione frontale superiore. Una piccola area sopra il solco cinguli, delimitata dietro dall'estremità posteriore del solco cinguli, e davanti da un piccolo solco, sulcus paracentrdlis, è chiamata lobulo circumcentrale, lobulo paracentrale, poiché corrisponde alla superficie mediale estremità superiori entrambe le circonvoluzioni centrali, che qui si incrociano.
Posteriormente al lobulo paracentralis è presente una superficie quadrangolare (il cosiddetto precuneo, precuneo), delimitato anteriormente dall'estremità del sulcus cinguli, inferiormente da un piccolo solco subparietdlis, e posteriormente da un profondo solco parietooccipitale. Il precuneo appartiene al lobo parietale. Dietro il precuneo si trova un'area nettamente separata della corteccia appartenente al lobo occipitale, il cuneo, cuneo, che è limitato davanti dal solco parietooccipitale e dietro sulcus calcarfnus, convergenti ad angolo. Inferiormente e posteriormente il cuneo è in contatto con il giro linguale. Il giro del cingolo si estende tra il solco cinguli e il solco del corpo calloso, giro del cingolo, che, attraverso l'istmo, istmo, continua nel giro parahippocampalis, terminando con un uncino, uncus. Il giro paraippocampale è limitato da un lato dal solco ippocampale, che circonda il tronco encefalico, e dall'altro dal solco collateralis e dalla sua continuazione anteriore, chiamato sulcus rhinalis. Istmo - il punto ristretto di transizione del giro cingolato nel paraippocampo, si trova dietro lo splenio corporis callosi, all'estremità del solco formato dalla fusione del solco parietooccipitale con il solco calcagno. Il giro cinguli, l'istmo e il giro parahippocampalis formano insieme il giro a volta, giro fornicato, che descrive un cerchio quasi completo, aperto solo in basso e davanti. Il giro a volta non è correlato a nessuno dei lobi del mantello.
Separando il bordo del solco dell'ippocampo, si può vedere una stretta striscia grigia frastagliata, che è un giro rudimentale del giro dentatus.
Struttura della corteccia cerebrale. La corteccia degli emisferi cerebrali nella sua tipologia principale è costituita da sei strati, che differiscono tra loro principalmente per la forma delle cellule nervose in essi incluse (Fig. 290): 1) lo strato molecolare si trova direttamente sotto la pia madre e contiene il rami terminali dei processi delle cellule nervose, intrecciati a rete; 2) lo strato granulare esterno, così chiamato perché contiene numerose piccole cellule granulari; 3) lo strato piramidale è costituito da cellule nervose piramidali di piccole e medie dimensioni; 4) lo strato granulare interno è composto, proprio come lo strato esterno, da piccole cellule - granelli; 5) lo strato gangliare contiene grandi cellule piramidali(cellule di Betz); 6) uno strato di cellule polimorfiche confina con la sostanza bianca. Di questi 6 strati, quelli inferiori (5° e 6°) costituiscono prevalentemente l'inizio delle vie efferenti; in particolare lo strato 5 è costituito da cellule piramidali di Betz, i cui assoni costituiscono il sistema piramidale. Gli strati intermedi (3° e 4°) sono associati principalmente a vie afferenti, e quelli superiori (1a e 2a) appartengono alle vie associative della corteccia. Il tipo di corteccia a sei strati viene modificato in varie aree sia in termini di spessore e disposizione degli strati, sia di composizione delle cellule (di cui si parlerà in dettaglio nel corso di istologia).
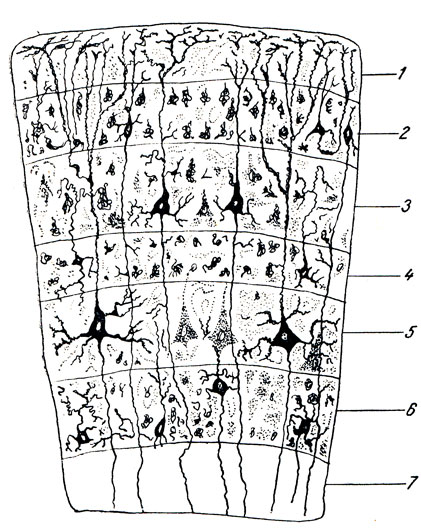
Cervello olfattivo (rinencefalo)
Cervello olfattivo, rinencefalo(Fig. 291, 292), è filogeneticamente la parte più antica del proencefalo, sorta in connessione con il recettore olfattivo, quando il proencefalo non era ancora diventato un organo di comportamento animale. Pertanto, tutti i suoi componenti lo sono varie parti analizzatore olfattivo(il concetto di analizzatore.
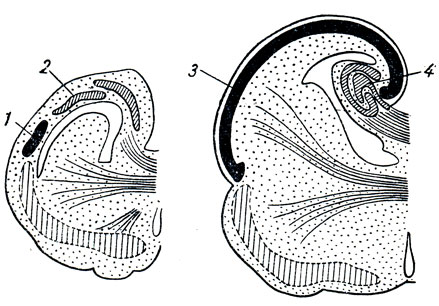
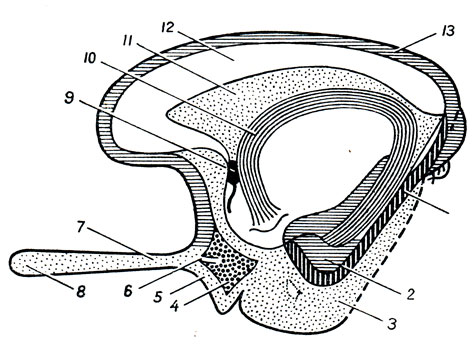
Nei pesci, quasi tutto il prosencefalo è l'organo dell'olfatto. Con lo sviluppo della nuova corteccia, che si osserva nei mammiferi e nell'uomo, si sviluppa una nuova parte del prosencefalo (neencefalo): il mantello, il pallio. Ma anche il mantello attraversa un lungo percorso di sviluppo e contiene tre parti di date filogenetiche diverse. Parti più vecchie:
1. Archipallium, parte del lobo temporale. Inizialmente, questa sezione si trovava sulla superficie laterale dell'emisfero, ma in seguito, sotto l'influenza del neopallio notevolmente ingrandito, si raggomitolò in una formazione a forma di salsiccia - l'ippocampo, o corno di Ammon, e si spostò medialmente nella cavità dell'emisfero. il ventricolo laterale del telencefalo sotto forma di sporgenza del suo corno inferiore. Il corno di Ammon è ricoperto di corteccia antica, archicorteccia.
2. Paleopallium, una piccola area della corteccia sulla superficie ventrale del lobo frontale, situata vicino al bulbus olfactorius e ricoperta da vecchia corteccia, paleocorteccia.
3. Neopallium, un nuovo mantello, nella cui corteccia apparve la neocorteccia centri superiori odore: le estremità corticali dell'analizzatore. Questo è un uncus, che fa parte del giro a volta.
Di conseguenza, il cervello olfattivo umano contiene una serie di strutture di varia origine, che può essere topograficamente suddiviso in due sezioni. La sezione periferica è il lobo olfattivo, lobus olfactorius, che si riferisce ad una serie di formazioni che si trovano alla base del cervello: 1) bulbus olfactorius; 2) tratto olfattivo; 3) trigonum olfactonum e 4) substantia perforata anteriore. Dipartimento centrale- queste sono le circonvoluzioni del cervello: 1) il giro del cavalluccio marino, l'ippocampo; 2) giro dentato, giro dentatus; 3) giro a volta, gyrus fornicatus, con la parte anteriore situata vicino al polo temporale - il gancio, uncus.
Ventricoli laterali
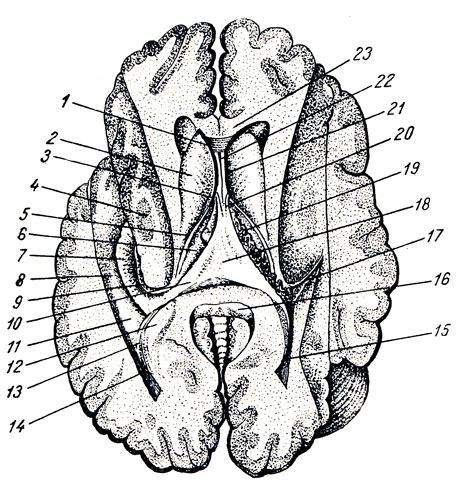
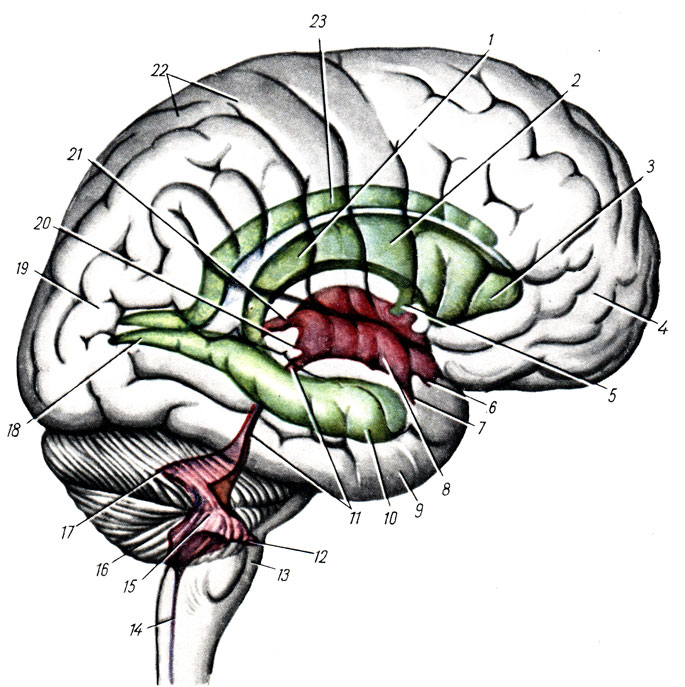
Negli emisferi, come residuo delle cavità originarie di entrambe le vescicole del telencefalo, si trovano al di sotto del livello del corpo calloso simmetricamente ai lati della linea mediana due ventricoli laterali, ventricoli laterali, separato dalla superficie superolaterale degli emisferi dall'intero spessore del midollo. La cavità di ciascun ventricolo laterale (vedi Fig. 294) corrisponde alla forma dell'emisfero: inizia nel lobo frontale sotto forma di un lato curvo verso il basso e laterale corno anteriore, cornu anterius, da qui si estende attraverso la regione del lobo parietale chiamata parte centrale, pars centralis, e poi a livello del bordo posteriore del corpo calloso si abbassa e avanza nello spessore del lobo temporale nella forma corno inferiore, cornu inferius, dove finisce. Nel punto in cui discende la cavità ventricolare, emana una proiezione posteriormente nel lobo occipitale - corno posteriore, cornu posterio.
La parete mediale del corno anteriore è formata dal setto pellucido, che separa il corno anteriore dallo stesso corno dell'altro emisfero (vedi Fig. 293). La parete laterale e in parte la parte inferiore del corno anteriore sono occupate dall'eminenza grigia, dalla testa del nucleo caudato, dai caput nuclei caudati e la parete superiore è formata da fibre del corpo calloso. Anche il tetto della parte centrale e più stretta del ventricolo laterale è costituito da fibre del corpo calloso, mentre il fondo è costituito dalla continuazione del nucleo caudato, dei nuclei dei corpi caudati e di parte della superficie superiore del talamo visivo. Il corno dorsale è circondato da uno strato di fibre nervose bianche originate dal corpo calloso, il cosiddetto tapetum (tappeto); sulla sua parete mediale è presente una cresta evidente: uno sperone di uccello, calcar avis, formato dalla depressione dal lato del sulcus calcarinus, situato sulla superficie mediale dell'emisfero. La parete superolaterale del corno inferiore è formata da un tapetum, che è la continuazione della stessa formazione che circonda il corno posteriore. Sul lato mediale, sulla parete superiore, è presente una parte assottigliata del nucleo caudato, piegata verso il basso e anteriormente - cauda nuclei caudati.
Lungo la parete mediale del corno inferiore per tutta la sua lunghezza è presente un rilievo bianco - Corno di Ammon, cornu Ammonis, o cavalluccio marino, ippocampo, che si forma a causa della depressione del solco ippocampi che taglia profondamente verso l'esterno. L'estremità anteriore dell'ippocampo è divisa da scanalature in numerosi piccoli tubercoli. Lungo il bordo mediale del corno di Ammon è presente una cosiddetta frangia, fimbria hippocampi, che rappresenta una continuazione del peduncolo dell'arco (crus fornicis). Alla base del corno inferiore è presente una cresta, eminentia collaterdlis, che nasce da una rientranza esterna al solco omonimo. Sul lato mediale del ventricolo laterale, un tessuto molle sporge nella sua parte centrale e nel corno inferiore. meningi, formando in questo luogo un plesso coroideo, il plesso chorioideus ventrulculi lateralis. Il plesso è ricoperto di epitelio, che rappresenta il residuo della parete mediale non sviluppata del ventricolo. Il plesso chorioideus ventriculi lateralis è il margine laterale del corpo della chorioidea ventriculi tertii.
Nodi o nuclei basali o centrali, emisferi
Oltre alla corteccia grigia sulla superficie dell'emisfero, nel suo spessore sono presenti anche accumuli di materia grigia, chiamati basale, centrale, O sottocorticale, nuclei e costituiscono quella che in breve viene chiamata "sottocorteccia". A differenza della corteccia, che ha la struttura dei centri schermo, i nuclei sottocorticali hanno la struttura dei centri nucleari. Esistono tre gruppi di nuclei sottocorticali: corpo striato, claustro e corpo amigdaloideo (Fig. 296, 297).
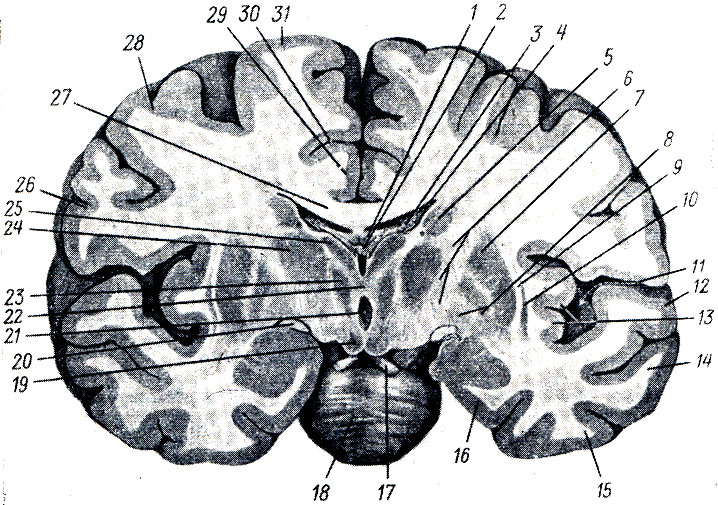
1. Corpo striato, striato, è costituito da due parti non completamente separate l'una dall'altra: il nucleo caudatus e il nucleo lentiformis.
UN. Nucleo caudato, nucleo caudato, si trova sopra e medialmente al nucleo lentiforme, separato da quest'ultimo da uno strato di sostanza bianca chiamato borsa interna, capsula interna. La parte anteriore ispessita del nucleo caudato, la sua testa, caput nuclei caudati, forma la parete laterale del corno anteriore del ventricolo laterale, mentre la sezione assottigliata posteriore del nucleo caudato, corpus et cauda nuclei caudati, si allunga, come abbiamo già visto, indietro lungo il fondo della parte centrale del ventricolo laterale; la cauda si avvolge parete superiore corno inferiore. Sul lato mediale, il nucleo caudato è adiacente al talamo ottico, separato da esso da una striscia di sostanza bianca, stria terminalis. Anteriormente e inferiormente, la testa del nucleo caudato raggiunge la substantia perforata anterior, dove si collega con il nucleo lentiformis (con la parte di quest'ultimo detta put amen). Oltre a questa ampia connessione di entrambi i nuclei sul lato ventrale, sono presenti anche sottili strisce di sostanza grigia intervallate da fasci bianchi della capsula interna. Servivano come motivo per il nome: striato, corpo striato (vedi Fig. 296).
B. Nucleo lentiforme, nucleo lenticolare, si trova lateralmente al nucleo caudato e al talamo visivo, separato da essi dalla capsula interna. In una sezione orizzontale dell'emisfero, la superficie mediale del nucleo lenticolare, rivolta verso la borsa interna, ha forma di angolo con l'apice rivolto verso il centro; il lato anteriore dell'angolo è parallelo al corpo caudato e il lato posteriore è parallelo al talamo visivo. La superficie laterale è leggermente convessa e si affaccia sul lato laterale dell'emisfero nella regione dell'insula. Anteriormente e ventralmente, come già indicato, il nucleo lenticolare si fonde con la testa del nucleo caudato. Nella sezione frontale il nucleo lenticolare ha la forma di un cuneo, il cui apice è rivolto verso la faccia mediale e la base è rivolta verso la faccia laterale. Il nucleo lenticolare è diviso in tre segmenti da due strati bianchi paralleli, lamine midollari, di cui quella laterale, di colore grigio scuro, è chiamata conchiglia, putamen(buccia), e i due mediali, più leggeri, sono chiamati collettivamente globus pallidus, globo pallido(vedi Fig. 296).
Differente già nell'aspetto macroscopico, il globo pallido ha anche una struttura istologica diversa dalle altre parti dello striato. Filogeneticamente, il globus pallidus è una formazione più antica (paleostriatum) del putamen e del nucleo caudatus (neostriatum).
Alla luce di tutte queste caratteristiche, il globo pallido è attualmente classificato in una speciale unità morfologica chiamata pallido, mentre la designazione striato è riservata solo al putamen e al nucleo caudatus. Di conseguenza, il termine “nucleo lenticolare” perde il suo significato precedente e può essere utilizzato solo in senso puramente topografico, e invece del precedente nome corpo striato, vengono chiamati nucleo caudato e lenticolare sistema striopallidale. Il sistema striopallidale è la parte principale del sistema extrapiramidale (vedi sotto) e inoltre è il centro regolatore più alto delle funzioni autonomiche in relazione alla termoregolazione e metabolismo dei carboidrati, dominante su centri vegetativi simili nell'ipotalamo.
2. Claustrum, recinzione, rappresenta una sottile lamina di materia grigia, situata nella regione dell'insula, tra questa e il putamen (vedi Fig. 297). È separata da quest'ultima da uno strato di sostanza bianca, capsula esterna, e dalla corteccia insulare da uno strato simile chiamato capsula extrema.
3. Corpo amigdaloideo, amigdala , o epistriatum, si trova sotto il putamen all'estremità anteriore del lobo temporale. Il corpo amigdaloideo appartiene apparentemente ai centri olfattivi sottocorticali. Termina con un fascio di fibre provenienti dal lobo olfattivo e dalla substantia perforata anteriore, segnalato nella descrizione del tubercolo visivo denominato stria terminalis (vedi Fig. 297).
Sostanza bianca degli emisferi
Tutto lo spazio tra la materia grigia della corteccia cerebrale e i nodi di maglia è occupato materia bianca. Consiste in grande quantità fibre nervose che corrono in direzioni diverse e formano le vie del telencefalo. Fibre nervose possono essere suddivisi in tre sistemi: 1) associazione, 2) commissurali e 3) fibre di proiezione.
UN. Fibre associative(Fig. 298) sono collegati tra loro varie aree corteccia dello stesso emisfero. Si dividono in brevi e lunghi. Le fibre corte, fibrae arcudtae cerebri, collegano le circonvoluzioni vicine sotto forma di fasci arcuati. Le fibre di associazione lunghe collegano aree della corteccia più distanti tra loro. Esistono diversi fasci di fibre di questo tipo. Cingulum, cintura, un fascio di fibre che passa attraverso il giro fornicatus, collega varie parti della corteccia del giro cinguli sia tra loro che con le vicine convoluzioni della superficie mediale dell'emisfero. Il lobo frontale è collegato al lobo parietale inferiore, al lobo occipitale e alla parte posteriore del lobo temporale attraverso il fasciculus longitudindlis superiore. I lobi temporale e occipitale sono collegati tra loro attraverso il fascicolo longitudindlis inferiore. Infine, la superficie orbitale del lobo frontale è collegata al polo temporale dal cosiddetto fascicolo uncinato, fasciculus uncindtus.
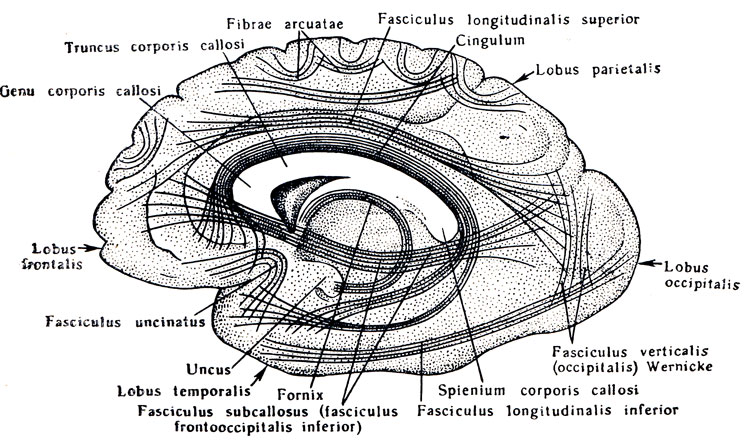
B. Fibre commissurali, che fanno parte delle cosiddette commissure cerebrali, o commissure, collegano le parti simmetriche di entrambi gli emisferi. La più grande commissura cerebrale è il corpo calloso, corpo calloso, che collega le parti di entrambi gli emisferi appartenenti al neencefalo.
Due commessure cerebrali, commissura anteriore e commissura fornicis, di dimensioni molto più piccole, appartengono al rinencefalo e collegano: commissura anteriore - lobi olfattivi ed entrambi i giri paraippocampali, commissura fornicis - corna di Ammon.
IN. Fibre di proiezione Collegano la corteccia cerebrale in parte con il talamo e i corpi geniculati, in parte con le parti sottostanti del sistema nervoso centrale fino al midollo spinale compreso. Alcune di queste fibre conducono le eccitazioni in modo centripeto, verso la corteccia, mentre altre, al contrario, in modo centrifugo.
Le fibre di proiezione nella sostanza bianca dell'emisfero più vicino alla corteccia formano la cosiddetta corona radiata, corona radidta, e quindi la parte principale di esse converge nella borsa interna, di cui sopra. La borsa interna, cdpsula interna, come già detto, è uno strato di sostanza bianca compreso tra il nucleo lentiforme da un lato e il nucleo caudato e il talamo ottico dall'altro. In una sezione frontale del cervello, la borsa interna appare come una striscia bianca obliqua che continua nel peduncolo cerebrale. In una sezione orizzontale, appare sotto forma di un angolo, aperto lateralmente (vedi Fig. 297), per cui la parte anteriore si distingue nella capsula interna, crus anterius capsulae internae - tra il nucleo caudato e la metà anteriore superficie interna nucleo lentiformis, parte posteriore, crus posterius, tra il tubercolo ottico e la metà posteriore del nucleo lentiforme, e il ginocchio, genu capsulae internae, situato nel punto di flesso tra entrambe le parti della borsa interna. Le fibre di proiezione possono essere divise in base alla loro lunghezza seguenti sistemi, a partire dal più lungo:
1. Trdctus corticospinale(pyramidalis), conduce impulsi motori volitivi ai muscoli del tronco e degli arti. A partire dalle cellule piramidali della corteccia della parte media e superiore del giro centrale anteriore e del lobulo paracentralis, fibre percorso piramidale vanno come parte della corona radiata, e poi passano attraverso la capsula interna, occupando i due terzi anteriori della sua parte posteriore, e le fibre per arto superiore andare davanti alle fibre per arto inferiore. Successivamente passano attraverso il peduncolo cerebrale, crus cerebri, e da lì attraverso il ponte nel midollo allungato.
2. Trdctus corticonucleare, che conducono percorsi ai nuclei motori dei nervi cerebrali. Partendo dalle cellule piramidali della corteccia della parte inferiore del giro centrale anteriore, passano attraverso il ginocchio della borsa interna e attraverso il peduncolo cerebrale, quindi entrano nel ponte e, spostandosi dall'altro lato, terminano nei nuclei motori del lato opposto, formando una decussazione. Una piccola parte delle fibre termina senza incrociarsi.
Poiché tutte le fibre motorie sono raccolte in un piccolo spazio nella borsa interna (il ginocchio e i due terzi anteriori della gamba posteriore), se vengono danneggiate in questo punto, si verifica una paralisi unilaterale (emiplegia) del lato opposto del corpo è osservato.
3. Trdctus corticopontini, vie dalla corteccia cerebrale ai nuclei pontini. Provengono dalla corteccia del lobo frontale (tractus frontopontinus), occipitale (tractus occipitopontinus), temporale (tractus temporopontino) e parietale (tractus parietopontinus). Come continuazione di queste vie, le fibre dei peduncoli cerebellari medi vanno dai nuclei pontini al cervelletto. Utilizzando questi percorsi, la corteccia cerebrale ha un effetto inibitorio e regolatore sull'attività del cervelletto.
4. Fascicoli talamocorticali e corticotaldmici, fibre dal talamo alla corteccia e di ritorno dalla corteccia al talamo. Tra le fibre provenienti dal talamo visivo è necessario notare la cosiddetta via tegmentale, che è la parte finale della via sensoriale diretta al centro del senso cutaneo nella circonvoluzione centrale posteriore. Emergendo dal nucleo laterale del talamo ottico, le fibre di questa via passano attraverso la parte posteriore della capsula interna, dietro il tratto piramidale. Questo luogo era chiamato “chiasma sensibile”, poiché qui passano altre vie sensoriali, ovvero il fascio ottico di Graziole, radiatio optica, che va dal corpo geniculatum laterale e pulvinar del talamo visivo al centro visivo nella corteccia del lobo occipitale , poi il fascio uditivo, radiatio acustica , che si estende dal corpo geniculatum mediale e dal tubercolo inferiore del quadrigemino alle circonvoluzioni di Heschl nel lobo temporale, dove si trova il centro dell'udito. I fasci visivi e uditivi occupano la posizione più posteriore nella parte posteriore della capsula interna.
Il corpo umano è un sistema molto complesso controllato da un computer incredibilmente potente: il cervello. Invia segnali a tutti gli organi e all'intero corpo, è responsabile di ciò che sentiamo, di come percepiamo il mondo e come interagiamo con esso. Sono passati secoli di evoluzione e le persone non sono ancora riuscite a comprendere appieno l'intero meccanismo di funzionamento del cervello. Lui è il principale parte integrale Sistema nervoso centrale.
Brevemente sulla cosa principale
La struttura del cervello umano è determinata dalle sue funzioni. Il cervello è coperto da tre tipi di membrane e in totale è costituito da 25 miliardi di neuroni, collettivamente chiamati materia grigia. Il cervello di una donna pesa leggermente meno di quello di un uomo, ma ciò è dovuto al processo evolutivo, non al grado di sviluppo. In media, la sua massa raggiunge il 2% della massa dell'intero corpo. Livello sviluppo mentale non dipende né dalle dimensioni del cervello né dalle dimensioni del corpo.
Struttura della corteccia cerebrale
Innanzitutto dovremmo parlare della corteccia. Il suo spessore è di 3 mm, copre le parti principali del cervello. La corteccia ha un aspetto molto struttura complessa, è composto da sei strati orizzontali che variano per dimensioni, densità, ecc. Ha anche il suo funzioni specifiche. Alcuni dei suoi lobi sono responsabili dell'olfatto, del tatto, della parola, ecc.
Struttura del cervello
Questo sistema complesso ha diversi dipartimenti, ciascuno con le proprie funzioni. Si distinguono i seguenti dipartimenti principali:
- L'ultimo rappresenta circa l'80% della massa dell'intero cervello.
- Intermedio: ecco il talamo e l'ipotalamo, che sono responsabili della connessione di una persona con l'ambiente.
- Posteriore: è costituito dal ponte e dal cervelletto, le cui funzioni sono strettamente correlate.
- Quello centrale è il dipartimento più “povero” in termini di funzioni.
- Oblongata: direttamente collegata al midollo spinale.
Inoltre, il cervello può anche essere suddiviso nelle seguenti sezioni:
- emisferi cerebrali;
- tronco;
- cervelletto.
Cervello finito
Questo reparto è probabilmente il più complesso sia per struttura che per funzione. Si compone di due sezioni: gli emisferi sinistro e destro, separati da una fessura. All'interno del solco, a loro volta, si trovano il fornice e il corpo calloso, che uniscono gli emisferi. Il telencefalo è la parte più funzionale dell'intero sistema.
L'emisfero sinistro è responsabile del pensiero astratto, mentre l'emisfero destro è responsabile del pensiero concreto. Inoltre, il telencefalo è responsabile della percezione vocale, emotiva e analitica, ottenendo informazioni su ambiente e altro ancora.
Diencefalo
Il talamo è direttamente responsabile della comunicazione con il mondo esterno, reagisce agli stimoli e trasmette informazioni su di essi agli emisferi. L'ipotalamo regola le funzioni autonomiche e interagisce con il sistema nervoso. Sotto di essa si trova la ghiandola pituitaria, le cui funzioni comprendono la regolazione del sonno e della veglia, il metabolismo e il controllo della temperatura corporea.
rombencefalo
Le funzioni del cervelletto e del ponte sono strettamente correlate. La struttura del cervello aiuta a capire come avviene la “comunicazione” tra i reparti. Dietro il ponte si trova il cervelletto, che funge in parte da conduttore. Il cervelletto è costituito da sostanza grigia e bianca ed è responsabile del coordinamento dei movimenti.
Mesencefalo
La gamma di funzioni di questo dipartimento è piccola, ma sono comunque molto importanti. La struttura del midollo spinale umano è tale da essere direttamente collegato al cervello attraverso il midollo medio. Trasporta impulsi causati da stimoli uditivi e visivi. Inoltre, è responsabile della cosiddetta visione nascosta e dell'orientamento del corpo verso il rumore. 
Midollo
È direttamente collegato al midollo spinale. La struttura di queste due parti del sistema nervoso ha molto in comune. Ecco qui materia bianca, che funge da canale di comunicazione che passa dal midollo spinale al cervello.
Conchiglie
Il cervello è ricoperto da diversi strati di membrane.
- La pia madre interagisce direttamente con il cervello; copre tutte le circonvoluzioni e i solchi. Inoltre, si ramifica nel cervello stesso e lo nutre.
- La membrana aracnoidea è la più sottile; entra in contatto con le circonvoluzioni, ma non le riempie.
- La dura è costituita da tessuto molto denso ed è collegata al cranio. Lo spazio tra l'aracnoide e la dura madre è pieno di liquido sieroso.

- Molti propagandisti immagine sana Nella vita dicono che l'alcol uccide le cellule cerebrali, ma in realtà questa è una bugia.
- Un giorno, un soldato ungherese fu colpito da un proiettile e il suo lobo frontale fu gravemente danneggiato. Sopravvisse, ma non riuscì a dormire per il resto della sua vita.
- Esiste un malinteso molto comune secondo cui le persone usano solo una piccola parte del loro cervello. Questo è fondamentalmente sbagliato. Il nostro cervello lavora 24 ore su 24 e a pieno regime.
- C'è anche un'opinione secondo cui le persone con un lato creativo più sviluppato ne hanno uno più sviluppato emisfero destro e le persone con pensiero analitico hanno una sinistra più sviluppata. Questo non è del tutto vero, infatti sono sviluppati allo stesso modo, solo che di conseguenza si è più attivi.
conclusioni
La struttura del cervello è molto complessa. Naturalmente, i dipartimenti e le funzioni di cui sopra rappresentano solo la conoscenza di base di questo sistema. In effetti, nel cervello ci sono molti più elementi e le loro connessioni sono molto più complesse. Processi lunghi l'evoluzione, durante la quale le funzioni del cervello, le sue dimensioni e la sua forma sono cambiate, ha portato al fatto che il cervello è diventato il “computer” più potente sulla Terra.
Nessuna macchina ha capacità così serie, nessun dispositivo può far fronte ai compiti assegnati così rapidamente. Si tratta di un sistema così complesso che, anche in un secolo di incredibili progressi in tutti i campi della conoscenza, le persone non sono state in grado di studiare a fondo il cervello, e nessuno può dire con certezza che ci riusciranno mai.